|
|
|
|
|
|
|
|
|
|
|
Not a few years ago, while driving through the pastoral oak woodlands of California's Gold Rush country en route to a couple of sensational fossil plant-bearing districts not far from The Biggest Little City In The world--Reno, Nevada--I knew that I needed to start the expedition off right by first visiting the very place where it had all begun, where I had initially learned of the superior localities I wanted to spend a few days examining from the undeniable doyen of Nevada paleobotany himself, the late paleobotanist Howard Schorn (passed in 2013), former collections manager of fossil plants at the University California Museum of Paleontology, he who had been among the first scientists to sytematically collect leaves, seeds, needles, fascicles, and cones from the places I intended to explore: the middle Miocene Pyramid and Chloropagus Formations of western Nevada, which produce 41 and 68 species of fossil plants, respectively. Once I'd finally found my way back to a long abandoned but once fantastically productive leaf-bearing quarry, having bushwhacked through thick tangled stands of manzanita and tight copses of overgrown scrub oak, I began to recollect back several years earlier to an early August field trip with Howard to this very spot in the western foothills of California's Sierra Nevada, Ione Basin, in the vicinity of Ione, Amador County, to help recover leaf impressions from an especially rich locality I had informally named Lygodium Gulch--an appropriate reference to the rather common remains of an extinct species of climbing fern found there--Lydodium kaulfusi, specifically, which is also known from locally spectacular occurrences in the early to middle Eocene Green River and middle Eocene Bridger Formations of Wyoming, Utah and Colorado, plentiful Polypodiopsida specimens whose closest modern-day counterpart is Lygodium palmatum, endemic to southern New England southward through the Appalachians into the southern states. It was a site I had discovered around 1994 during exploratory reconnaissance in the paleobotanically and geologically remarkable middle Eocene Ione Formation, approximately 48 million years old, an early Cenozoic sedimentary sequence consisting primarily of fluviatile, estuarine, deltaic, marine, and marsh deposits that not only contain locally prodigious quantities of often well preserved semi-tropical vegetation, mainly bountiful leaves (numerous palm fronds found, as well) stained an aesthetically pleasing reddish brown on a brilliant white shale matrix partially composed of detrital feldspathic constituents, but also world-class horizons of high value ceramic clays, pure silica sands, and lignites that bear--in a global perspective--extraordinarily rare commercial-grade quantities of montan wax, used in such diverse applications as fruit coating to phonograph records; along with the Ione Formation example, Amsdorf Germany is the only other place on Earth that mines commercial quality montan wax-yielding coal. Howard and I were there as an advance guard, as it were, to collect fossil specimens intended for a National Science Foundation project, secured by Howard, paleobotanist Jack A. Wolfe, and several other scientists to help determine the paleoelevation and paleoenvinment of the ancestral Eocene Sierra Nevada area. Along with supplemental leaf material collected from the middle Eocene auriferous gravels higher up the western slopes of the Sierra Nevada (fluviatile and lacustrine accumulations usually considered a proximal paleohydrologic manifestation of the same watercourses that supplied detritus to the distal Ione Formation), exposed at several abandoned hydraulic gold mines that date from roughly 1855 to 1889, Dr. Wolfe wanted to run the Ione Formation fossils through his vaunted CLAMP system--that is to say, the Climate Leaf Analysis Multivariate Program, an accurate methodology that analyzes 31 different physiognomic parameters of fossil leaves (among them, lobing, margin characteristics, size, apex, and base) to help determine the past elevation and environment of a given fossil plant locality. In order to efficiently secure at Lygodium Gulch the necessary quantity of paleobotanical material within our lone week of allotted field activity (Wolfe required at least 20 different leaf morphologies to analyze with confident precision a given paleo-flora), Howard eventually made a decisive command decision--one firmly settled upon, I might add, after we'd spent an arduously exhausting first full day whacking repeatedly into the unyielding ground in search of fossil leaves; we'd uncovered several excellent specimens amenable to Dr. Wolf's CLAMP analysis, but the going was brutally tedious, working as we were with repetitive backbreaking technique, pick and shovel brute force paleontology under a sizzling Sierran foothill summer sun that had topped 100 degrees before noon: Howard chose to hire a local backhoe operator to come on out to our fossil locality and entrench the lithified, highly compacted leaf-bearing sedimentary rocks, so that in the pleasant aftermath of the bobcat's excavations all we then had to do was sit around on the spoils piles of that trench and happily crack apart the fluviatile shales and sandstones at our leisure in the early and mid hours of successive mornings, eventually amassing numerous boxes brimmed with finely detailed middle Eocene leaves that Howard took back to the archival paleobotany collections at the University California Museum of Paleontology. Today, of course, the roughly 48 million year old semi-tropical leaves of Lygodium Gulch lie within what botanists call the Ione Chaparral, a unique association of plants that inhabits Ione Basin, western Amador and extreme northern Calaveras Counties, western foothills of California's Sierra Nevada. It's a botanical province found nowhere else on Earth, restricted solely to oxisoils that erode from the harshly acidic and highly mineralized middle Eocene Ione Formation. Typical botanic components of an Ione Chaparral include the following specimens: Ione manzanita--Arctostaphylos myrtifolia, the most characteristic member of the Ione Chaparral, a rare and endangered low shrub, wholly captivating with its striking diminutive dark green leaves and minute bell-like flowers (they typically bloom from December through February), which grows nowhere else in the wild except in scattered disjunct populations atop the acidic, toxically mineralized oxisoils of the middle Eocene Ione Formation within a narrow 19.5 mile north-south trending geographic corridor of California's Ione Basin; white leaf manzanita--Arctostaphylos viscida; common manzanita--Arctostaphylos manzanita; Hellers's manzanita (a natural hybrid between Ione manzanita and white leaf manzanita)--Arctostaphylos helleri; Ione buckwheat--Eriogonum apricum (a rare and federally protected botanic specimen that grows nowhere in the wild, except in California's Ione Basin); Irish Hill buckwheat--Eriogonum apricum var. prostratum (another rare and endangered buckwheat found only in the wild at Ione Basin, California); coyote brush--Baccharis pilularis; chamise--Adenostoma fasiculatum; yerba santa (sacred herb--here's one to ruminate on, to put in your proverbial pipe and smoke: the native Chumash Indians of California used yerba santa in steam baths to treat hemorrhoids, and its flavinoid sterubin might eventually help alleviate Alzheimer's symtomatology)--Eriodycton californicum; interior live oak--Quercus wislizenii; California scrub oak--Quercus berberidifolia; and California foothill pine (an older, now seldom-used designation is the Digger pine)--Pinus sabiniana, in addition to various native grasses, and the cryptogam mosses and lichens--Cladonia cervicornis ssp. verticillata, primarily, a lichen restricted to mineral-rich subsurface areas where Ione manzanita dominates (the lichen is not present in adjacent regions where abundance organic litter accumulates). Why Ione manzanita is confined to such a restricted geographic district in California remains a mystery. Even though the Ione Formation can be traced for some 200 miles throughout the western foothills of the Sierra Nevada, from roughly Oroville south to Friant, east of Fresno, the Ione manzanita grows atop Ione exposures only along that narrow 19.5 mile swath of the Ione Basin; perhaps maritime breezes that rush through the Carquinez Strait, San Francisco Bay region, situated 90 miles southwest of the Ione Basin (as the crow flies), help moderate summer temperatures enough to maintain viable stands of Ione manzanita within such a geographically restricted habitat. A companion idea centers around those abundant lichens present in Ione Formation soils. As advanced by a researcher in 1995, a working hypothesis postulates that after periodic episodes of fire--a necessary requirement for Ione manzanita seeds to germinate--the lichen thallus releases substantial quantities of aluminum, an element the lichens efficiently assimilate from the highly mineralized middle Eocene Ione Formation; that critical increase in aluminum concentration inhibits most other chaparral plants from establishing early footholds in the Ione Basin, because seedlings of Ione manzanita preferentially tolerate such usually toxic levels of aluminum while other young chaparral plants do not. To bolster his idea, the specialist adduced evidence that as mature specimens, Ione manzanita and the common white leaf manzanita both flourish atop low pH (acidic) Ione Formation oxisoils with equally high concentrations of aluminum in their plant cells, demonstrating to his own satisfaction at least that based on levels of aluminum uptake alone, no special edaphic differentiation exists between the two manzanita species; he then concluded that any natural adaptive advantage conferred upon Ione manzanita probably stems from an ability to better survive the seedling stage in an extraordinarily challenging environment exponentially enriched in toxic aluminum released by lichens after fires. Along with studying the Ione manzanita and associated Ione chaparral, collecting paleobotanical remains in the middle Eocene Ione Formation certainly gave me an increased appreciation of changing environments over the course of deep geologic time. Originally deposited in floodplains near the eastern shores of a vast inland Eocene sea that had once covered most of California's Great Central Valley 48 million year ago, the Lygodium Gulch locality contains abundant semi-tropical vegetation comprised entirely of leaves with smooth margins, lacking serrations or lobes, whose numerous species could undoubtedly thrive today alongside their modern analogs in proximity to southern Florida's mangrove swamps, where annual precipitation regularly approaches 45 inches, including substantial showers delivered during the summer months. Today, the Ione Formation fossils occur in the lower western foothills of the Sierra Nevada, in the neighborhood of Ione, California, within a Mediterranean meteorological regime characterized by hot dry summers lacking effective rainfall, where showers usually relegated to winter through spring drop about 21 inches of precipitation per year, a dramatic alteration of prevailing weather patterns, rainfall totals, and geography that directly influences the differences in vegetational representation so obvious between that which lived here 48 million years ago and the modern chaparral community dominated by manzanita and chamise and scrub oak and interior live oak, and California foothill pine. While we split shales that had been dumped around the perimeter of the trench created by our bobcat operator, Howard likely identified rather quickly my insatiable interest in such paleoenvironmental considerations. And so by and by he suggested that I should get out to western Nevada, around Reno, to investigate two additional fantastic paleobotanical localities that provide important information on drastically different environments over the course of millions of years: With meticulous attention to detail, he described the middle Miocene Pyramid and Chloropagus Formations, 15.6 and 14.8 to 13.3 million years old, respectively, where nicely preserved leaves, seeds, needles, fascicles, cones and cone scales prove that today's botanically depauperate Great Basin Desert scene around Reno not only once held lush deciduous hardwood trees similar to types now found in southeastern Tennessee, but also--a couple of million years later--a giant sequoia forest community reminiscent of Calaveras Big Trees State Park, western slopes of California's Sierra Nevada, not far from Gold Rush Country. Seems that Howard had actually accompanied legendary paleobotanist Daniel I. Axelod to the sites, helping to secure some of the earliest scientific collections from the Pyramid and Chloropagus Formations, fabulous fossil material now housed at the University California Museum of Paleontology. That was how it all started. Howard had graciously provided me with invaluable paleobotanical information on a couple of Nevada Neogene sites--floristically fascinating fossil places that had instantly captured my imagination. And so it came to pass that almost a year after that Ione Basin expedition with Howard, after I'd attended to all that which needed attention in the interim (a lot of mundane particulars happily interspersed with periodic spurts of paleontological inquisitiveness), I finally got around to getting out into the field once again; I loaded up my trusty four wheel drive mechanism with needed supplies and important scientific references pertaining to paleobotany, botany, and geology and lit out for the territory. First up on my paleobotany itinerary was a visit to the middle Miocene Pyramid Formation in the neighborhood of Reno, Nevada. Today, the 15.6 million year old middle Miocene Pyramid Formation fossil locality (older of the two sites I explored) lies near Reno at roughly 4,800 feet elevation in an arid to semi-arid ecotone situated between Great Basin Desert and Basin Sage botanic communities, where it receives a sparse five to eight inches of effective precipitation per year, with more than fifty percent of it falling as snow. At the fossil site today, mean temperatures for each month of the year are (in Fahrenheit): January--30 degrees; February--33 degrees; March--40 degrees; April--50 degrees; May--60 degrees; June--70 degrees; July--74 degrees; August--72 degrees; September--63 degrees; October--50 degrees; November--38 degrees; and December--34 degrees. The effective growing season lasts on average some 133 days. By contrast, based on the known environmental preferences of modern-day counterparts of plants preserved in the fossil flora, elevations during deposition of the middle Miocene Pyramid Formation 15.6 million years ago could not have been any higher than 1,800 to 2,000 feet. To support the vegetational varieties identified from the Pyramid site, the ancestral Reno region necessarily received at minimum 35 inches of annual precipitation, and 35 percent of that total needed to fall during May through August (in order to account for several exotic deciduous hardwoods in the fossil flora). In general, winter months were much milder and summers cooler than conditions experienced at the fossil locality today. Growing season lasted an expected 177 days, a figure calculated from approximated average monthly temperature conditions for middle Miocene Pyramid times (all degrees in Fahrenheit): January--43 degrees; February--47 degrees; March--52 degrees; April--58 degrees; May--64 degrees; June--69 degrees; July--70 degrees; August--70 degrees; September--67 degrees; October--57 degrees; November-- 48 degrees; and December--45 degrees. In the immediate proximity of the Pyramid Formation paleobotanical bonanza, plant life is typical of a Great Basin Desert-Basin Sage transition zone: Sparse shadscale-type vegetation occupies low-lying areas, while increasing elevations above 5,000 feet support juniper (Juniperus utahensis), mountain mahogany (Cercocarpus ledifolius), basin sage (Artemisia tridentata), rabbit bush (Chrysothamnus rauseosus), desert peach (Prunus andersonii), plateau gooseberry (Ribes velutinum), and bitterbrush (Purshia tridentata). Rare moist areas reveal wild rose (Rosa gymnocarpa), cottonwood (Populus fremontii), several species of willows (Salix sp.), chokecherry (Prunus cemissa), dogwood (Ccrnus californica), and hairy horsebrush (Tetradymia glabrata). Higher mountain slopes underlain by geologically altered igneous rocks contain occasional specimens of Jeffrey pine (Pinus jeffreyi). Now go back some 15.6 million years to that same geographic area and you'd observe quite a different botanic scene: A dominantly deciduous hardwood community of maple (Acer), alder (Alnus), birch (Betula), Kentucky yellowood (Cladastris), ash (Fraxinus), American hophornbean (Ostrya), sycamore (Platanus), willow (Salix), Chinese scholar tree (Pterocarya), and poplar (Populus) contribute to lush forested lakeshores, floodplains, and riparian places, while bald cypress (Taxodium) and a now extinct species of willow oak (Quercus simulata) grow in swampy areas. Along drier well-drained volcanic slopes above the basin of deposition, you'd take in a mixed conifer-deciduous association of fir (Abies), incense cedar (Calocedrus), spruce (Picea), pine (Pinus), mountain hemlock (Tsuga), chokecherry (Prunus), mountain ash (Sorbus), and wild rose (Rosa), with such broadleafed evergreen sclerophylls as madrone (Arbutus), mountain mahogany (Cercocarpus), giant chiquapin (Chrysolepis), and tanbark oak (Lithocarpus) also well represented. The middle Miocene Pyramid Formation fossil plants were discovered in 1951 by entomologist Ira La Rivers of the University Nevada Reno during field investigations not far from Reno; he expeditiously relayed the information to paleobotanist Daniel I. Axelrod, who initially collected a modest supply of specimens there in 1952 with funds from the Carnegie Institute of Washington. In 1957, Geologist George S. McJannet first mapped the fossil-producing area and masterfully pinpointed a type locality for what he named the Pyramid Formation. Later on down the line, in the early 1960s, Axelrod secured a National Science Foundation Grant to continue scientific studies of the fossil flora, and that's the general time period when paleobotanist Howard Schorn, collections manager of fossil plants at the University California Museum of Paleontology, assisted with the Pyramid plant project, helping Axelrod collect additional leaf, seed, and needle specimens from a single spot in the proximity of Reno, Nevada. In order to more efficiently collect enough fossil specimens from the poorly exposed sedimentary beds to accommodate a scientific analysis, Axelrod and Schorn soon opened up the locality with a caterpillar. In all, they amassed some 1,414 plant remains from McJannet's middle Miocene Pyramid Formation, more than enough fossil material to help Axelrod complete a monographic study of the Pyramid Formation plants. Axelrod's paleobotanical locality lies in middle member C of McJannet's Pyramid Formation (for ease of stratigraphic evaluation, conveniently divided into three mappable units), a 250 foot-thick section that includes: bone-white leaf-bearing diatomite, a sedimentary rock type composed primarily of diatoms, miscroscopic single-celled photosynthesizing kinds of algae that constructed intricately designed frustules composed of silica; subordinate, organically barren shales and sandstones derived from local volcanic plugs; and unfossiliferous reworked swirls of rhyolitic and dacite sands produced by turbidity currents. Youngest member A, overlying the plant-yielding horizon, is 220 feet of scoriaceous dark-gray to black basalt with minor tuff and breccia, and the oldest unit C consists of yellow-brown basalt, tuff, breccia, with a conglomerate of rhyolite and dacite clasts at the base (770 feet thick). The middle Miocene Pyramid Formation is but one of several igneous-metamorphic-sedimentary sequences exposed in the general area, and the stratigraphy on display there is quite fascinating. Oldest rocks that outcrop around the fossil locality constitute a roughly 80 million year old late Cretaceous basement complex comprised of: granitic aplite and pegmatite dikes; hornblende biotite quartz diorite; a roof pendant of andalusite-feldspar-biotite schist; quartz plagioclase amphibolite; greenish gray hornblende; and plagioclase cataclasite--all of undetermined thickness. Above the Mesozoic material lies a significant unconformity in the local geologic record, a mysterious 55 million year old interval of depositional quiescence when prolonged erosive processes ultimately erased all local traces of the succeeding Paleocene and Eocene Epochs; probably at least part of this geologic time frame coincides with fluvial generation of the world-famous fossil plant-bearing middle to upper Eocene auriferous gravels and the stratigraphically equivalent Ione Formation, western foothills of the Sierra Nevada, by rivers that transected the ancient paleodivide, situated in Eocene times between present-day Battle Mountain and Austin, Nevada. Lying directly atop the unconformity is the Tule Peak Formation, a 25 to 23 million year old upper Oligocene to lower Miocene volcanic accumulation approximately 3,740 feet thick, consisting of, in descending order of geologic age: welded rhyolite; white pumiceos crystal tuff; massive white pumice lapilli; reddish-brown welded vitric crystal rhyolite tuff with local basalt flows; light to dark welded rhyolite tuff that outcrops with a blocky fracture; collapsed pumice lapilli (lenticulite); and roughly 10 to 20 feet of arkose and breccia at the base. Next youngest local geologic rock unit is the 23 million year old lower Miocene Sutcliff Formation, another exclusively igneous unit some 1,140 feet thick that includes, from top to bottom (youngest to oldest): hard, massive to platy welded rhyolite breccias with associated lenticulites (collapsed pumice lapilli); lavender to gray welded biotite rhyolite tuff, usually massive; and resistant well-indurated rhyolite tuffs that outcrop in distinctive massive fashion. Protruding above the Sutcliff Formation in unconformable geologic relations are several geomorphological features earth scientist call stocks, or volcanic plugs composed of massive reddish-brown quartz dacite that solidified from magmas inside volcanoes around 20 to 21 million years ago; they now form picturesque peakes and slopes, against which the 15.6 million year old leaf-bearing diatomites of the middle Miocene Pyramid Formation lap unconformably, signifying that several million years of erosion occurred between the last active volcano and lacustrine deposition of the overlying Pyramid Formation diatomite. Another unconformity separates the 15.6 million year old Pyramid Formation from the overlying 1,920 foot thick middle Miocene Chloropagus Formation, roughly 13.3 to 14.8 million years old. Although not exposed in the immediate vicinity of the Pyramid Formation leaf locality, elsewhere near Reno the unit indeed provide paleobotany adventurers with leaves, seeds, needles, fascicles, cones, and cone scales from some 68 species of plants. Typical Chloropagus outcrops possess a dominant igneous component, chiefly green, black, dark-brown and reddish vesicular basalt and andestie flows, interbedded with occasional volcanic tuff breccias and agglomerates; intercalcating sedimentary constituents include water-laid tuff, tuffaceous sandstone, and locally fossiliferous siliceous fissile shales roughly three and a quarter to six and a half feet thick. Next youngest member of the local straigraphic column, positioned unconformably above the middle Miocene Chloropagus Formation, is a mainly igneous middle Miocene section approximately 420 feet thick, dated by ratiometric methods at 12.7 million years old. Its three unnamed mappable subunits, members, include from youngest to oldest: flow-banded dacite vitrophyre with columnar jointed, collapsed pumice altered to perlite; buff coarse-grained poorly indurated dacite tuff, tuffaceous sandstones, and conglomerate; and dark-gray to black latite associated with tuff breccia and black collapsed pumice. Resting unconformably atop the Tertiary Period exposures is a 515 foot thick aggregate of Quaternary Pleistocene-age sedimentary-volcanic material approximately 100,000 years old, probably representing at least four different Ice Age depositional events: (1)--an upper (youngest) sequence consists of playa silts, sands, and gravels that grade into lake clays; (2)--a distinctive unit of relatively well exposed clay, sand and gravel that correlates stratigraphically with distant sedimentary evidences of ancient Lake Lahontan--one of the largest bodies of fresh water ever to exist in Pleistocene age inorthwestern Nevada; (3)--a dark green to black olivine basalt flow; and (4)--a basal (oldest) unit of poorly sorted, weakly consolidated sands. The Pyramid Formation fossils accumulated through current transport in a relatively small basin bordered by extinct volcanic plugs and active volcanism. Along with macroscopic vegetation, microscopic diatom species accumulated in a shallow cool clear lake saturated for prolonged periods with dissolved silica, stimulating repeated prodigious algal reproduction cycles that eventually deposited a diatomite bed about 200 feet thick. 88 percent of the 1,414 Pyramid specimens belong to deciduous hardwoods. Geographic distribution analyses demonstrate that eleven of the 41 fossil plants are allied with kinds that now grow in the eastern US; 24 Pyramid species presently reside in the western US; and five others show close affinity with endemics in China, Mexico, and East Asia. In a generalized correlative trend, the Pyramid remains also reveal a greater relationship with Miocene fossil floras from the Columbia Plateau district of Oregon, Washington, and Idaho than they do with identical-age Miocene Nevadan paleobotanical sites. Modern areas that support vegetation similar to associations that grew in western Nevada during deposition of the middle Miocene Pyramid Formation include the Yuba River drainage, western foothills of California's Sierra Nevada, in the transitional ecotone situated between a mixed conifer forest and a broadleaf sclerophyll community (chaparral), where several varieties closely allied to fossil types found in the 15.6 million year old middle Miocene Pyramid Formation now grow, among them--Ponderosa pine (Pinus ponderosa), Douglas-fir (Pseudotsuga menziesii), incense cedar (Calocedrus masonii), Pacific madrone (Arbutus menziesii), chinquapin (Chyrsolepis), tanbark oak (Lithocarpus), Western sycamore (Platanus racemosa), box elder (Acer negundo), bigleaf maple (Acer macrophyllum), Oregon ash (Fraxinus oregona) dusky willow (Salix melanopsis), sandbar willow (Salix exigua), and wild rose (Rosa); and around Metcalf Bottoms, southeast of Townsend, Tennessee, a floodplain region that hosts a mesic deciduous hardwood forest of plant species similar to kinds recovered from the middle Miocene Pyramid Formation, including species of maple (Acer), buckeye (Aesculus), alder (Alnus), birch (Betula), Kentucky yellowtree (Cladastris), ash (Fraxinus), American hophornbean (Ostrya), and chokecherry (Prunus). Most common 21 fossil species found in the middle Miocene Pyramid Formation include, in decreasing order of relative abundance: (1) leaves and acorns from a presumably extinct oak called Quercus simulata (possibly related to the living willow oak Quercus phellos, southeastern United States, or even the silver oak Quercus hypoleucoides of southeastern Arizona, southwestern New Mexico, and northern Mexico); (2) leaves of Betula thor (fossil counterpart to the living paper birch Betula papyrifera); (3) leaves and cones from an alder Alnus latahensis (Miocene counterpart of Alnus maritima, the seaside alder or brook alder); 4) leaves from a second species of alder Alnus spokanensis (the fossil equivalent of the modern speckled alder Alnus incana subspecies rugosa); samaras and leaves from a maple Acer chaneyii (the Miocene variety of today's sugar maple Acer saccharum); (5) leaves of a buckeye Aesculus preglabra (paleotanical genus-species for the living Ohio buckeye Aesculus glabra); (6) seeds, needles, and twigs from a pine Pinus ponderosoides (Miocene variety of the living Ponderosa pine--also called western yellow-pine--Pinus ponderosa); (7) leaves that belong to Populus bonhamii (the Miocene variety of Populus balsamifera, the balsam poplar); (8) branchlets from a swamp cypress Taxodium oregonensis (the Miocene genus-species of Taxodium distichum, the modern bald/swamp cypress); (9) leaves of Lithocarpus nevadensis (the fossil equivalent to the extant tanbark-oak Notholithocarpus densiflorus); (10) leaves from Platanus bendirei (this is what paleobotanists call fossil structures that are identical to today's California sycamore Platanus racemosa, also referred to as western sycamore); (11) leaves and samaras of Acer oreganianum (paleobotanical genus-species for the living bigleaf maple Acer macrophyllum); (12) leaves belonging to Prunus chaneyii (the fossil equivalent to the extant Western chokecherry Prunus virginiana var. demissa); (13) leaves from Acer glabroides (a species analogous to the living red maple Acer rubrum); (14) winged seeds and cone scales that come from Abies laticarpus (the Miocene genus-species for the modern Abies magnifica var. shastaensis--the living Shasta red fir); (15) leaves of Salix pelviga (this is the paleobotanical genus-species for the extant dusky willow Salix melanopsis); (16) leaves derived fromAlnus pyramidensis (paleobotanical name given to fossil structures identical to the extant alder Alnus cremastogyne); (17) leaves that belong to Betula vera (the Miocene equivalent to the modern yellow birch--also called golden birch or swamp birch--Betula lutea; (18) leaves of Salix churchillensis (the Miocene binomial name for fossil remains that show close relationship with today's sandbar willow Salix exigua subspecies exigua); (19) leaves that belong to Arbutus idahoensis (according to paleobotanist Daniel I. Axelrod, this is a presumably extinct species of madrone whose closest modern-day affinity is possibly with the Texas madrone Arbutus xalapensis); (20) foilage and seeds from Calocedrus masonii (name paleobotanists apply to fossil structures identified as indistinguishable from counterparts produced by the extant Calocedrus decurrens, the incense cedar--not a true cedar, of course); (21) leaves from Cladrastis lariverisii (the fossil analog for the living Kentucky yellowwood Cladastris lutea. The remaining 20 Pryamid Formation species are rather rarely encountered, having been described in the scientific literature from but three or fewer fossil specimens. Here is the complete list of the 41 fossil plant species recovered from the middle Miocene Pyramid Formation, 15.6 million years old. Number that follows each description is the total number of specimens of that species recovered from the Pyramid Formation: 1) Abies laticarpus (winged seeds and cone scales)--The Miocene genus-species for the modern Abies magnifica var. shastaensis--the living Shasta red fir which ranges from 1,400–2,700 meters (4,600–8,900 ft) elevation in Shasta, Siskiyou and Trinity Counties, northwest California, through southwest Oregon. 9. 2) Picea sonomensis (winged seeds)--Fossil name paleobotanists apply to the extant Picea breweriana, the Brewer spruce, one of the rarest trees in North America, which now grows in the wild only in the Klamath Mountains of southwest Oregon and northwest California at elevations from 1,000–2,700 m (3,300–8,900 ft). DNA genetic analyses determined that the Brewer spruce lies at the base of the Picea clade, suggesting that spruces originated in North America. 1. 3) Pinus ponderosoides (twigs, needles, seeds)--Miocene variety of the living Ponderosa pine (also called western yellow-pine), Pinus ponderosa that in modern natural habitat ranges as various subspecies variants southward through British Columbia into 16 western US states at elevations from 100–2,700 m (330–8,860 ft). 32. 4) Pseudotsuga sonomensis (seeds and branchlets)--Paleobotanical Miocene counterpart for the extant Douglas-fir, Pseudotsuga menziesii, a member of the Pinus (pine) family (not a true fir, hemlock or spruce), naturalized in British Columbia, Canada, southward through Washington, Oregon, California, and Nevada, USA. Elevations typically range from 550m -2,900m (1,800 to 9,500 ft), though some river bottom populations live at 800 to 900 feet. 2. 5) Tsuga mertensioides (seeds)--The fossil variety of today's Tsuga mertensiana, mountain hemlock, which presently ranges from the Kenai Peninsula, Alaska, southward to northern Tulare County, California, usually at elevations from: sea level to 1,000 m (3,300 ft)--Alaska; 1,600 to 2,300 m (5,200 to 7,500 ft)--Cascades in Oregon; and 2,500 to 3,050 m (8,200 to 10,010 ft)--Sierra Nevada, California. 1. 6) Calocedrus masonii (foliage and seeds)--Name paleobotanists apply to fossil structures identified as indistinguishable from counterparts produced by the extant Calocedrus decurrens, the incense cedar (not a true cedar, of course). Present-day native geographic range: central-southwestern Oregon; most of California; extreme western Nevada; and northern Baja California, Mexico. Invariably grows at elevations from 50–2,900 m (160–9,510 ft). 4. 7) Taxodium oregonensis (branchlets)--The Miocene genus-species of Taxodium distichum, the modern bald (swamp) cypress, now native to the warm, humid southeastern United States with annual precipitation levels of 760 mm or 30 inches (eastern Texas) to 1,630 mm--64 inches (Gulf Coast). 27. 8) Populus bonhamii (leaves)--The Miocene variety of Populus balsamifera, the balsam poplar (northernmost hardwood in North America), now native to most of Canada and the following US states: Alaska, northeastern Oregon, central and northwestern Idaho, scattered in five isolated areas in Montana, Wyoming, north to central Colorado, northwestern Nebraska, western and northeastern South Dakota, southeastern and northern North Dakota, northern half of Minnesota, northern Iowa, Wisconsin, Michigan, northeastern Illinois, northwestern Indiana, northeastern Ohio, Pennsylvania, northeastern West Virginia, western Maryland, northeastern Connecticut, New York, Vermont, northern half of New Hampshire, and Maine. 30. 9) Salix boisienis (leaves)--Fossil genus-species for the living Salix scouleriana (Scouler's willow), now endemic to western to central Canada and the following US states: mainly southeastern Alaska, Washinton, Oregon, California (primarily northwestern and the Sierra Nevada area), western Nevada, Idaho, western Montana, western South Dakota, Wyoming, Colorado, Arizona, and New Mexico. 1. 10) Salix churchillensis (leaves)--The Miocene binomial name for fossil remains that show close relationship with today's sandbar willow (coyote willow) Salix exigua subspecies exigua, which currently ranges southward from southern Canada through eastern Washington, eastern Oregon, eastern California, Idaho, Nevada, southwestern Montana, western Wyoming, Utah, Colorado, Arizona, New Mexico, western Texas, and northern Mexico. 5. 11) Salix inquirenda (leaves)--The Miocene equivalent of today's Salix lasiandra, the Pacific willow, which attains a wide geographic distribution, ranging from Alaska southward through western to central Canada, Washington, Oregon, California, Idaho, Montana, Wyoming, Colorado, Arizona, and New Mexico. 2. 12) Salix pelviga (leaves)--This is the paleobotanical genus-species for the extant dusky willow Salix melanopsis, now native to British Columbia and Alberta southward to California and Colorado, preferring moist habitats along riverbanks and subalpine mountain meadows. 8. 13) Alnus latahensis (leaves and cones)--The Miocene counterpart to the living Alnus maritima, the seaside alder or brook alder now found naturally in Oklahoma, Georgia, Maryland, and Delaware; it is the only North American member of the genus Alnus that blooms during autumn--all other alders bloom in spring. 242. 14) Alnus pyramidensis (leaves--named in honor of having been first identified from the middle Miocene Pyramid Formation)--Paleobotanical name given to fossil structures that are identical to the extant alder Alnus cremastogyne, now endemic to southwestern China along stream banks and forested mountainsides at elevations from 500 - 3,000 meters (1.640 to 9,842 feet). 6. 15) Alnus spokanensis (leaves)--The fossil equivalent of the modern speckled alder Alnus incana subspecies rugosa, which today ranges in natural habit from Canada southward through northern North Dakota, Minnesota, Wisconsin, Michigan, northeastern Iowa, northern Illinois, northern Indiana, northern Ohio, Pennsylvania, western Maryland, eastern West Virginia, New York, Connecticut, Rhode Island, Massachusetts, Vermont, New Hampshire, and Maine. 161 16) Betula ashleyii (leaves)--Bionmial name that paleobotanists give to the extant water birch (red birch) Betula fontinalis (synomymous with Betula occidentalis), which in natural geographic habitat ranges southward through southern Canada to form scattered occurrences in Washington, Oregon, California, Idaho, Nevada, Arizona, New Mexico, Colorado, Montana, Wyoming, North Dakota, South Dakota, and western Nebraska. 1. 17) Betula thor (leaves)--The fossil counterpart to the living paper birch (also called American white birch) Betula papyrifera, now native to almost all of Canada and most of Alaska, southward to parts of the following US states: Washington, Oregon, Idaho, Montana, North Dakota, South Dakota, Nebraska, Colorado (southernmost occurrence), Minnesota, Wisconsin, Michigan, Illinois, Iowa, Indiana, Ohio, New York, West Virginia, North Carolina, Pennsylvania, Connecticut, Massachusetts, and Rhode Island. It also occurs throughout all of Connecticut, Vermont, New Hampshire, and Maine. 286. 18) Betula vera (leaves)--The Miocene equivalent to the modern yellow birch (also called golden birch or swamp birch) Betula lutea (synonymous with Betula alleghaniensis). Prefers cooler environmental condisitions, usually in swamps, stream banks, deep woods, and north facing slopes from southeastern Canada southward into northeastern Minnesota, Wisconsin, northeastern Iowa, northernmost Illinois, northern Indiana, northeastern Ohio, Michigan, Pennsylvania, New York, Vermont, New Hampshire, Maine, Connecticut, Massachusetts, Rhode Island, northern New Jersey, northern Maryland, West Virginia, western half of Virginia, Kentucky, Tennessee, western North Carolina, northwestern South Carolina, and northeastern Georgia. 7. 19) Ostryea oregoniana (leaves)---This is the paleobotanical designation for today's Ostrya virginiana, the American hophornbeam. Has wide geographic distribution:: Occurs in Nova Scottia west to Manitoba and then southward into Minnesota, Iowa, Missouria, Arkansas, Lousiana, eastern Kansas, eastern Oklahoma, eastern Nebraska, eastern North Dakota, eastern Texas, western Wyoming, southeastern and western South Dakota, Wisconsin, Illininoid, Michigan, Indiana, Ohio, Pennsylvania, New York, Vermont, New Hampshire, Maine, Rhode Island, Massachusetts, Connecticut, Maryland, New Jersey, West Virginia, Virginia, Kentucky, Tennessee, North Carolina, South Carolina, Florida, Alabama, and Mississippi. 2. 20) Pterocarya mixta (leaves)--The Miocene analog to the living Chinese wingnut Pterocarya stenoptera, a species commonly cultivated horticulturally for shade and landscaping; it is endemic to the following Chinese provinces on riverbanks or forested mountains from around sea level to 1500 m. (4,921 feet): Anhui, Fujian, Gansu, Guangdong, Guangxi, Guizhou, Hainan, Hebei, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Liaoning, Shaanxi, Shandong, Shanxi, Sichuan, Yunnan, and Zhejiang. Disjunct populations also found in Taiwan, Japan, and Korea. 2. 21) Chrysolepis sonomensis (leaves)--The paleobotanical name for the living giant chinquapin Chrysolepis chrysophylla, endemic to southern Washington southward through western Oregon and west-central California, favoring climax forests dominated by redwood, white fir, western hemlock, and Shasta red fir. 3. 22) Lithocarpus nevadensis (leaves)--The fossil equivalent to the extant tanbark-oak Notholithocarpus densiflorus, presently native to California and southern Oregon. Despite its common name and the fact that it does produce acorns, it is nevertheless not a true oak. 26. 23) Quercus simulata (leaves and acorns)--A presumably extinct species whose fossil leaves and acorns match no known modern oak. Several paleobotanists suggest that it's possibly allied with the living willow oak Quercus phellos, native to the southeastern United States, or even the silver oak Quercus hypoleucoides, presently found in southeastern Arizona, southwestern New Mexico, and northern Mexico. 378. 24) Platanus bendirei (leaves)--This is what paleobotanists call fossil structures that are identical to today's California sycamore Platanus racemosa (also referred to as western sycamore), a common riparian specimen that ranges mainly from western and eastern California--northernmost occurrence is in Humbolt County, with scattered patches in the Sierra Nevada--south to northern Baja California Mexico. 25. 25) Ribes standordianum (leaves)--A Miocene genus-species whose modern-day counterpart is Ribes nevadense, the Sierra currant (or, mountain pink currant), mainly native to riparian and forest habitats in California's mountainous regions (Sierra Nevada, Peninsular Ranges, Transverse Ranges, and Klamath Mountains, for example) at elevations between 3,000–10,000 feet (910–3,050 meters); also distributed in western Nevada and Oregon. 1. 26) Cercocarpus ovatifolius (leaves)--This is the fossil analog to the living Catalina mahogany/island mountain mahogany Cercocarpus betuloides var. blancheae, endemic to coastal southern California, where it's confined in natural development to scattered occurrences in Los Angeles County, Ventura County, Santa Barbara County, San Luis Obisbo County, and the Channel Islands. 2. 27) Prunus chaneyii (leaves)--The fossil equivalent to the extant Western chokecherry Prunus virginiana var. demissa, presently native to northern, southern and central California, preferring in natural habitat rocky slopes at elevations from 0-8200 feet. 28) Rosa shornii (leaves)--Named in honor of paleobotanist Howard Schorn, former collections manager of fossil plants in the University California Museum of Paleontology, Berkeley, California; the Miocene analog to the modern California wild rose Rosa californica now native to California (notably, coast and foothills up to elevations of 6,000 feet), Oregon, and northern Baja California Mexico. 3. 29) Sorbus mcjannetii (leaves)--The Miocene counterpart of today's whitebeam/mountain ash Sorbus scopulina, whose native habitat ranges from central and southern Alaska, southward through western Canada to Washington (scattered occurrences), northern Idaho, western Montana, northwestern Wyoming, northern Utah (with scattered occurrences in central southern areas), western South Dakota, northeastern Nevada, Colorado, New Mexico, northeastern Arizona, and northern California (one isolated occurrence in central California). 3. 30) Cladrastis lariverisii (leaves)--The fossil analog for the living Kentucky yellowwood Cladastris lutea (synonymous with Cladrastis kentukea), which today grows in the wild only in Kentucky, Tennessee, Arkansas, Mississippi, Indiana, western Oklahoma, Alabama, western North Carolina, western Virginia, northern Georgia, and southern Missouri. 4. 31) Sophoroa spokanensis (leaves)--Paleobotanical name given to the modern Chinese scholar tree (also called Japanese pagoda tree) Styphnolobium japonicum (synonymous with Sophoroa japonica), native to China, Korea, and Japan where it prefers open country habitats at elevations from 300 and 1000 meters (984 to 3,280 feet). 1. 32) Acer chaneyii (leaves and samaras)--This is what paleobotanists call the Miocene variety of today's sugar maple Acer saccharum, a geographically widespread botanic specimen that in natural habitat grows in southeastern Canada, southward to Minnesota, eastern Iowa, Missouri, eastern Kansas, southeastern North Dakota, northeastern South Dakota, Wisconsin, Illinois, Michigan, Indiana, Ohio, Pennsylvania, New York, Vermont, New Hampshire, Maine, Connecticut, Massachusetts, Rhode Island, northern New Jersey, northern Maryland, western Virginia, West Virginia, Kentucky, Tennessee, western North Carolina (with scattered occurrences in the central part of the state), Georgia (two isolated areas in the north), and South Carolina (one isolated area in the northwestern portion of the state). 46. 33) Acer glabroides (leaves)--A fossil species analogous to the living red maple Acer rubrum, presently distributed in geographic range from southeastern Canada southward into the following US states: eastern Texas, eastern Oklahoma, Missouri, Minnesota, Michigan, Wisconsin, Indiana, southern Illinois, Ohio, Pennsylvania, New York, Connecticut, Vermont, New Hampshire, Maine, New Jersey, Maryland, Rhode Island, Massachusetts, West Virginia, Virginia, North Carolina, South Carolina, Georgia, Florida, Mississippi, Alabama, Louisiana, Arkansas, Tennessee, Kentucky. 11. 34) Acer negundoides (leaves)-- The binomial paleobonical name given to the fossil representative of the modern box elder Acer negundo, one of the most widespread species of tree in America, occurring naturally in every US state, except for Washington, Oregon, Maine, and Rhode Island. It's also present in south-central Canada and ranges as far south as Guatemala. 1. 35) Acer oreganianum (leaves and samaras)--Paleobotanical genus-species for the living bigleaf maple Acer macrophyllum, found primarily in southern Alaska, extreme western Canada, and then south through the coastal regions of Washington, Oregon,, and California (a disjunct population inhabits the western foothills of the Sierra Nevada) at elevations from sea level to 450 m (1,480 ft), with unusual exceptions to 1,200 m (3,900 feet). 21. 36) Acer scottiae (leaves)--The fossil name for the modern yellow-paint maple Acer pictum, an Asian species widespread in temperate forests across, China, Korea, Japan, Mongolia, and eastern Russia. 3. 37) Acer septilobatum (leaves)--A presumably extinct species of maple that shows some similarities to the extant bigleaf maple Acer macrophyllum, which lives in western coastal areas of the US and Canada (with a geographically separated occurrence in the western foothills of California's Sierra Nevada). 1. 38) Aesculus preglabra (leaves)--Paleobotanical genus-species for the living Ohio buckeye Aesculus glabra whose natural geographic distribution includes populations in Ohio, southern Michigan, eastern Pennsylvania, Tennessee, northern Alabama, Mississippi (one isolated spot near the Alabama border), central to northeastern Texas, eastern half of Oklahoma, northwestern Arkansas, western Kansas, Missouri, southern Iowa, Illinois, and Indiana. 34. 39) Arbutus idahoensis (leaves)--According to paleobotanist Daniel I. Axelrod, this is a presumably extinct species of madrone whose closest modern-day affinity is possibly with the Texas madrone Arbutus xalapensis, native to Central America, Mexico, and the southwestern United States (Texas and New Mexico). 5. 40) Arbutus matthesii (leaves)--The Miocene analog to the living Pacific madrone Arbutus menziesii, native primarily to coastal western North America, from southern Canada south to California--they are rare south of Santa Barbara County, and an isolated population inhabits the western foothills of the Sierra Nevada. 2 41) Fraxinus caudata (leaves)--The paleobotanical genus-species for the modern Oregon ash Fraxinus oregona (synonymous with Fraxinus latifolia), native to the US west coast from western Washington southward through western Oregon to northwest and central California; it prefers moist loose soils at elevations from sea level to 900 m (3,000 feet--in the north) to 1,700 m (5,600 feet--in California). 3. Part 2--A visit to the 13.3 to 14.8 million year old middle Miocene Chloropagus Formation. Next up, I spent a couple of days exploring the geologically younger middle Miocene Chloropagus Formation. At roughly 14.8 to 13.3 million years old, the Chloropagus fossil flora occurs in the vicinity of Reno, western Great Basin Desert, Nevada, and records a significant regional botanic transition, beginning about 15 million years ago, from a landscape previously characterized by deciduous hardwoods allied with today's eastern US Appalachian area to an environment that would have more closely resembled a modern mixed conifer forest along the western slopes of the Sierra Nevada, California. That's when exotic angiospermous hardwoods and bald cypress (Taxodium) underwent a rather rapid (geologically speaking) extirpation from mid Miocene western Nevada in response to decreased precipitation during the months of May through August; Chloropagus times received fully five inches less rain during late spring through summer than the Pyramid Formation plants only one and a half million years earlier. This famous paleobotanic crisis is excellently reflected in nine geologically stratified Chloropagus Formation florules that preserve in superior detail the traumatic transition. Perhaps mid Miocene East Antarctic ice sheet spreading, combined with Australia's separation from Antarctica and a concomitant widening of Drake's passage when Continental Drift also moved South America away from Antarctica, drove colder waters northward toward the tropical districts, strengthening high pressure in the southern United States and Mexico, especially in the ancestral west. Whatever the explanation, relic middle Miocene hardwood forests in Oregon and Washington managed to escape the severe botanic turnover until approximately two to three million years later, as the great drying trend moved inexorably northward. But of course, over the past 14 million years or so the area's vegetational trend has reflected increasing aridity. Occurring not far from Reno Nevada today, the middle Miocene fossil plant-bearing Chloropagus Formation outcrops at roughly 4,400 feet within the parched western reaches of the Great Basin Desert, receiving perhaps a little as five inches of rain per year, with most of it falling as snow during winter. In general, mean monthly temperatures at the fossil sites today show greater summer and winter extremes than the same seasonal weather conditions estimated for Chloropagus times (all readings in Fahrenheit): January--30 degrees; February--37 degrees; March--43 degrees; April--50 degrees; May--57 degrees; June--66 degrees; July--73 degrees; August--72 degrees; September--63 degrees; October--53 degrees; November--40 degrees; and December--32 degrees. Growing season in the neighborhood of the Chloropagus Formation fossil localities today is no more than four months. Yet, 14.8 to 13.3 million years ago, based on the known environmental preferences of modern counterparts of the fossil flora, the Reno-vicinity region within which the Chloropagus Formation accumulated stood at an elevation no higher than 2,400 feet and received, initially, an estimated 30-35 inches of precipitation annually--an amount that had dropped to 25 to 30 inches near the conclusion of sedimentary deposition, when May to August rainfall had all but disappeared. During the summer and winter months especially, middle Miocene Chloropagus climatic conditions would have been characterized by temperate temperature gradients decidedly more moderate than those observed today in the western Great Basin Desert (all figures in Fahrenheit): January--40 degrees; February--43 degrees; March--45 degrees; April--52 degrees; May--59 degrees; June--65 degrees; July 66 degrees; August--63 degrees; September--58 degrees; October--52 degrees; November--47 degrees; and December--42 degrees. The effective growing season likely lasted as long as seven months. Plants at the fossil sites today reflect the restrictive effects imposed by demanding Great Basin Desert meteorology. Now dominating a landscape that had once held luxuriant green forests of conifers is an association of specialized low spiny plants adapted to scant rainfall (often less than five inches per year) and extreme temperature fluctuations, where summer highs peak at 106 F. and winter lows can drop to minus 26 degrees Fahrenheit. Typical shrubs encountered today include sage (Artemesia); saltbrush (Atriplex); blackbrush (Coleogyne); fireweed (Kochia--an introduced non-native species, of course); siltbush (Grayia); and horsebrush (Tetradymia). The only trees present in the area are Fremont cottonwood (Populus fremontii) and various species of willow (Salix), invariably concentrated near rare water sources. This contrasts radically with the abundant and diverse Chloropagus vegetation present in the middle Miocene Reno region 14.8 to 13.3 million years ago, when a lush forested scene featured numerous species of conifers, deciduous trees and shrubs, broadleafed evergreen sclerophylls, and herbaceous perennials such as fir (Abies), spruce (Picea), pine (Pinus), larch (Larix), mountain hemlock (Tsuga), Lawson cypress, (Chamaecyparis), Chinese swamp cypress (Glyptostrobus), Douglas-fir (Pseudotsuga), juniper (Juniperus), giant sequoia (Sequoiadendron), maple (Acer), alder (alnus), birch (Betula), scarlet hawthorn (Crataegus), Chinese coffee tree (Gymnocladus), sycamore (Platanus), cottonwood (Populus), quaking aspen (Populus); extinct species of willow oak (Quercus simulata); zelkova (Zelkova); serviceberry (Amalanchier), moutain mahogany (Cercocarpus), ocean spray (Holodiscus); dogwood (Cornus); chokecherry (Prunus); buckthorn (Rhamnus); black locust (Robinia); wild rose (Rosa); highbush blueberry (Vaccinium); madrone (Arbutus), giant chinquapin (Chrysolepis); tanbark oak (Lithocarpus); Santa Cruz Island iron tree (Lyanothamnus--a species that today grows in the wild only on the Channel Islands off the coast of southern California); canyon live oak (Quercus); tobacco brush (Ceanothus); a brush-sized mountain mahogany (Cercocarpus); toyon (Heteromeles); Sierra laurel (Leucothoe); Island barberry (Mahonia); Tall Oregon grape (Mahonia); horsetail (Equisteum); broadleaf cattail (Typhus); and eagle fern (Pteridium). The middle Miocene Chloropagus Formation fossil plant bonanza was discovered in 1952 by University Nevada Reno Mackay School of Mines geologist/minerologist Dr. Vincent P. Gianella (1886-1983), who delivered a small batch of specimens he'd gathered to paleobotanist Dr. Daniel I. Axelrod, then teaching at the University California at Los Angeles (infamous UCLA--grin: This is the first and only time I will ever mention UCLA in writing; I am a University of Southern California devotee, of course: Enough said, I reckon). It's not clear, though, who actually secured the next Chloropagus finds; on that account, Axelrod is rather vague, noting only that several succeeding collections came while working out stratigraphic relationships in the fossiliferous district. He doesn't directly "confess" to personally gathering the specimens, though. It should of course be noted at this juncture that in a rather fortuitous earth science connection, professor Gianella served for a number of years as a de facto fossil-finding stringer for Axelrod, who benefitted greatly from Gianella's original field investigations. On not a few occasions, for example, Dr Gianella graciously informed Axelrod of places in Nevada where identifiable fossil leaves could be found, significant paleobotanical spots that Axelrod would have never been aware of otherwise; the wonderful Virginia City-vicinity middle Miocene fossil leaf locality immediately comes to mind in this regard, a district that not only produces numerous identifiable leaves and seeds, but also many insects and occasional frog skeletons, as well. In 1969, geologist Harold Bonham with the Nevada Bureau of Mines forwarded more Chloropagus Formation fossil plant material to Axelrod, now a professor of botany and paleobotany at the University California Davis in northern California (he became a member of the faculty there in 1968). Axelrod accompanied Bonham to the new plant-bearing areas where they subsequently discovered four remarkable superposed fossil floras, each stacked one atop another in normal stratigraphic succession (that is to say, no unconformities to interrupt a continuous sedimentary column). That instantly re-invigorated the Chloropagus fossil project, and up until 1976 when he "officially" retired from UC Davis, Axelrod regularly took his students in paleobotany and paleoecology on field trips to the "stacked fossil plant place" to collect additional leaves, seeds, needles, fascicles, cones and cone scales, ultimately amassing some 5,295 specimens from nine localities. At one of those especially productive sites, paleobotanist Howard Schorn ably assisted collecting fossil plants for Ax--a well-known nickname for Daniel I. Axelrod, by the way, and in actual fact he actively encouraged its use; while at UC Davis, for example, he had a personalized license place that read PROF AX. And when it came time to assemble for publication his monograph on the middle Miocene Chloropagus plants, Axelrod enlisted Howard Schorn to photograph representative specimens to accompany the text and charts. All the fossil plants that Axelrod described in his scientific treatise came from minor shale intervals in the middle Miocene Chloropagus Formation; a potassium-argon radiometric reading from a basalt flow near the base (oldest level) of the geologic unit yielded a chronostratigraphic date of 14.8, while another radioactive isotope analysis (presumably K/Ar) from a glassy rhyolitic tuff close to the top (youngest horizon) produced an absolute age of 13.3 million years. In its generalized geologic aspect of outcropping, the Chloropagus is predominantly of volcanic origin, bearing locally prolific paleobotanical remains only within several discontinuous pale brown to black lacustrine shale beds, usually no more than three to seven feet thick (perhaps representing geologically short-lived lake or pond waters that had been impounded due to volcanism), that outcrop in traditionally difficult to spot gulches and gullies eroded into approximately 1,900 feet of reddish-brown to light greenish gray basalt and andesite flows (in individual flow beds 10 to 49 feet thick); adding to their considerable paleontological mystique, the richest fossil-bearing shales are impossible to trace outside of the nondescript, localized erosion channels within which they remain confined. In the three primary areas where fossil plant material occurs--appropriately labeled sites 1, 2, and 3--the middle Miocene Chloropagus Formation lies in varying stratigraphic relationship to other igneous rock units. Area 1 for example contains two 14.8 million year old fossil florules situated at the base (oldest horizon) of a Chloropgus Formation section nearly 300 feet thick; directly underlying the thinly bedded plant-bearing Chloropagus lacustrine shales is the middle Miocene Old Gregory Tuff, around 20 feet of rhyolite tuff and breccia dated at 15 million years old. Directly below the Old Gregory Tuff (in unconformable stratigraphic position) is the 22 million year old lower Miocene Alta andesite, probably a little over 300 feet thick, which in turn overlies an undisclosed thickness of welded tuffs belonging to the 28 to 23 million year old Olligocene to lower Miocene Hartford Hill Rhyolite, a once long-established geologic formation name that in the Carson City-Silver City area has been abandoned in favor of the following new rock units, from oldest to youngest: Mickey Pass Tuff; Lenihan Canyon Tuff; Nine Hill Tuff; and Eureka Canyon Tuff. Below the Hartford Hill Rhyolite, separated by a great unconformity that left no trace of the preceding Eocene and Paleocene Epochs, is a Mesozoic basement complex of unestablished thickness composed of Cretaceous-age hornblende biotite quartz diorite and granidiorite, underlain in turn by older Jurassic and Triassic Period metasediments that include dark hornfels, argillite, metamorphosed limestones and some metavolcanics. At paleobotanical locality 2, four famous superposed fossil localities occur in a 962 foot thick section of the Chloropagus Formation, which unconformably overlies the lower Miocene Alta Andesite, dated by radiometric isotope techniques at 22 million years old, a roughly 300 foot thick sequence of greenish-gray to dark green and dark gray hornblende and pyroxene andesite flows bearing subordinate breccias mottled with blotches of red and green. Area 3 provides three paleo-florules scattered within about 738 feet of Chloropagus Formation outcropping; one fossil plant zone occurs roughly 100 feet above the unconformably underlying 28 to 23 million year old Oligocene to lower Miocene Hartford Hill Rhyolite, while the remaining two plant-yielding localities--estimated at 13.3 million years of age--lie some 100 feet below the conformably overlying Kate Peak Peak Andestie, which consists of hornblende andesite flows, mudflows, breccias, tuffs, conglomerates and some diatomite beds, all convincingly dated through radiometric methodology at 13 million years old. Daniel I. Axelrod had the four superposed fossil localities of area 2 under scientific study for some three decades. He noted that each individual florule reflects changing vegetational associations directly influenced by repeated episodes of successional extrusive volcanism over the course of perhaps 100,000 years. The paleobotanical preservations thus provide an extraordinary opportunity to evaluate through a relatively "brief" period of geologic time the effects of volcanic activity on a middle Miocene geography roughly 14 million years ago. The oldest plant horizon yields a flora composed of fully 84 percent sclerophyllous evergreens, dominated by giant chinquapin (Chrysolepis), tanbark oak (Lithocarpus), and canyon live oak (Quercus); this association suggests a rocky well drained soil profile that developed on rather recent lava flows subjected to erosion. A conifer element comprised of fir (Abies), Lawson cypress (Chamaecyparis lawsiana), spruce (Picea), Douglas-fir (Pseudotsuga), and giant sequoia (Sequoiadendron) contributed but four percent of the total take, probably inhabiting patches of moister slopes nearby. Pond-lake components such as willow (Salix) and cottonwood (Populus) are also comparatively rare here, providing only four percent of the 343 identified specimens, reinforcing the interpretation that recently weathered well drained volcanic soils covered most of the terrain closest to paleobotanical accumulations. Lying only 10 feet above the oldest florule in successional stratigraphic relations is a three foot thick plant-bearing horizon of gray-black organic shale that bears a significant proportion (36 percent) of riparian plants, principally birch (Betula) cottonwood (Populus) and willow (Salix). Broadleaf evergreen sclerophylls such as madrone (Arbutus), mountain mahogany (Cercocarpus), and toyon (Heteromeles) contributed only 20 percent of the 227 total specimens studied; they were likely confined to drier sites at the edges of volcanic flows in a region of subdued relief. A large mixed conifer element (44 percent) and its associates suggest a relic old growth forest established near a pond or small lake, where fir (Abies), Lawson cypress (Chamaecyparis lawsiana), spruce (Picea), Pine (Pinus), Douglas-fir (Pseudotsuga), serviceberry (Amalanchier), chokecherry (Prunus), and mountain ash (Sorbus) regularly shed their botanic structures to the sedimentary record. Roughly 50 feet higher in the section at locality 2 is a third superposed florule. Here members of a rich mixed conifer forest community that probably lived on highly weathered volcanic terrain account for fully 67 percent of the 967 plant specimens studied. Regular gymnospermous elements included fir (Abies), Lawson cypress (Chamaecyparis lawsiana), spruce (Picea), Douglas-fir (Pseudotsuga), and giant sequoia (Sequoiadendron). Compared with their prominence in the oldest horizon, broadleaf sclerophylls such as madrone (Arbutus), mountain mahogany (Cercocarpus), toyon (Heteromeles), and canyon live oak (Quercus pollardiana) became greatly diminished during deposition of the third Chloropagus successional florule, forming only 19 percent of the total; the more xeric vegetation probably grew on less weathered soils than their forest counterparts, or perhaps at the edges of younger volcanic flows which provided access to increased sunlight and much drier environments. A pond-lake border association of birch (Betula), cottonwood (Populus) and willow (Salix) lived near lake level, accounting for 17 percent of the curated paleobotanical material. The youngest fossil zone at Chloropagus area 2 lies roughly 56 feet above the third florule. In numerical representation of fossil species, it is certainly the most prolific of the four superposed horizons, yielding 36 species of middle Miocene plants identified from 870 specimens. By this time, a conifer forest with subordinate associated taxa had become dominant (74 percent of the fossils examined), a diverse botanic community consisting of fir (Abies), Lawson cypress (Chamaecyparis lawsiana), spruce (Picea), Douglas-fir (Pseudotsuga), giant sequoia (Sequoiadendron), serviceberry (Amalanchier), birch (Betula), snowbrush ceanothus (Ceanothus), scarlet buckthorn (Crataegus), Sierra laurel (Leucothoe), black locust (Robinia), mountain ash (Sorbus), Tall Oregon grape (Mahonia), and western chokecherry (Prunus). The broadleaf sclerophyllous component of madrone (Arbutus), mountain mahogany (Cercocarpus), toyon (Heteromeles), Island barberry (Mahonia), oak (Quercus), and tanbark oak (Lithocarpus) underwent a reduction (nine percent of the total), occupying more exposed xeric sites on volcanic flows near the basin of deposition. Nearest the lake level lived a thriving deciduous plant association of (Betula), cottonwood (Populus), and willow (Salix) that contributed 17 percent of the fossil specimens known from the uppermost superposed Chloropagus zone. Today, four geographic areas support plant communities similar to those preserved in the middle Miocene Chloropagus Formation, Great Basin Desert, Nevada, including: (1) Calaveras Big Trees State Park, western foothills of California's Sierra Nevada, amidst a rich mixed conifer forest, where several conifers, sclerophylls, and shrubs closely allied with fossil types found in the 13.3 to 14.8 million year old Chloropagus Formation in the vicinity of Reno, Nevada, now grow--including: giant sequoia (Sequoiadendron), fir (Abies), pine (Pinus), Douglar-fir (Pseudotsuga), madrone (Arbutus), tanbark oak (Lithocarpus), canyon live oak (Quercus), serviceberry (Amalanchier), buckbrush (Ceanothus), black laurel (Leucothoe), Oregon grape (Mahonia), western chokecherry (Prunus), wild rose (Rosa), and willow (Salix); (2) Sugar Lake, Salmon Mountains--a branch of the Klamath Mountains--northwestern California, within the southern sector of the Miracle Mile, a world famous square mile of natural "real estate" that holds one of the most diverse conifer assemblages on Earth, including foxtail pine (Pinus balfouriana), whitebark pine (Pinus balfouriana), western white pine (Pinus monticola), Jeffrey pine (Pinus jeffreyi), ponderosa pine (Pinus ponderosa), lodgepole pine (Pinus contorta), sugar pine (Pinus lambertiana), white fir (Abies concolor), Shasta fir (Abies magnifica), subalpine fir (Abies lasiocarpa), Engelmann spruce (Picea engelmannii), Brewer spruce (Picea breweriana), mountain hemlock (Tsuga mertensiana), Douglas-fir (Pseudotsuga menziesii), Pacific yew (Taxus brevifolia), incense-cedar (Calocedrus decurrens), common juniper (Juniperus communis), and western juniper (Juniperus occidentalis)--upwards of 32 species of conifers have been identified from the Klamath Mountains region; (3) Along the eastern slopes of Cone Peak, Santa Lucia Range, California, where Shasta red fir (Abies magnifica), sugar pine (Pinus lambertiana), madrone (Arbutus), canyon live oak (Quercus), tanbark oak (Lithocarpus) , Oregon grape (Mahonia), mountain mahogany (Cercocarpus), dogwood (Cornus), toyon (Heteromoles), oceanspray (Holodiscus), pigeon berry (Amalanchier), western chokecherry (Prunus), wild rose (Rosa), and buckthorn (Rhamnus) now grow; and (4) Northern Idaho, where many conifers and shrubs allied with fossil kinds found in the 13.3 to 14.8 million year old Chloropagus Formation occur, including larch (Larix), mountain hemlock (Tsuga), fir (Abies), Douglas-fir (Pseudotsuga), western serviceberry (Amalanchier), scarlet hawthorn (Crataegus), oceanspray (Holodiscus), oregon grape (Mahonia), wild rose (Rosa), willow (Salix), western chokecherry (Prunus), and whitebeam (Sorbus). Here is the full list of fossil plant species identified from the middle Miocene Chloropagus Formation as exposed in the neighborhood of Reno, Nevada, Great Basin Desert. The nine numbers that follow the descriptions refer to the numbers of fossil specimens of that species found at each of the nine fossil localities, arranged in order of oldest plant-bearing horizon to youngest (left to right); in other words, first number refers to the oldest zone and the last number is the youngest of the nine horizona: 1) Chamaecyparis cordillerae (synonym of C. sierrae) (twigs, foliage)--The Miocene equivalent of today's Lawson cypress, Chamaecyparis lawsiana, now native to southwestern Oregon and the Klamath Mountains of northwest California at elevations from sea level to 4,900 feet (1,500 m). 3, 18, 3, 20, 42, 36, 3, 7, 0. 2) Abies nevadensis (winged seeds)--Paleobotanical counterpart to the extant Sakhalin fir Abies sachalinensis which today growns on Sakhalin island, southern Kurils (Russia), and northern Hokkaido (Japan) at elevations from sea level to 1,650 meters (5,413 feet), preferring some 1,500mm of precipitation annually (about 60 inches). 0, 0, 0, 0, 0, 1, 0, 0, 0. 3) Abies laticarpus (synonym of A. klamathensis) (winged seeds)--The Miocene analog to Abies magnifica var. shastaensis--the living Shasta red fir that in natural habitat ranges from 1,400–2,700 meters (4,600–8,900 ft) elevation in Shasta, Siskiyou and Trinity Counties--northwest California--through southwest Oregon. This species also occurs in the 15.6 million year old Pyramid Formation. 10, 2, 7, 3, 0, 66, 3, 1, 0. 4) Abies scherrii (cone scales)--The fossil genus-species of the modern Shasta Lucia fir Abies bracteata, now confined in natural habitat to the Santa Lucia Mountains near Big Sur, central California coast, where it grows at elevations from 2,000 to 5,000 feet (610 to 1,520 m). 1, 1, 0, 0, 3, 5, 0, 0, 0. 5) Larix fernleyii (winged seeds, twigs, and needles)--Name paleobotanists use to scientifically describe fossil plant structures that closely resemble those produced by the living subalpine larch Larix lyallii, whose natural habitat ranges from the Rocky Mountains of British Columbia, Alberta, Idaho, and Montana to an isolated population in Washington's Cascade Range, preferring elevations from 1,500 to 2,900 meters (4,900 to 9,500 feet). 0, 1, 0, 6, 3, 1, 0, 0, 0. 6) Picea magna (winged seeds)--A presumably extinct spruce, although a few paleobotanists suggest that it does resemble the living tigertail spruce Picea torano, now native to Japan. 0, 220, 6, 12, 0, 0, 3, 52, 0. 7) Picea sonomensis (winged seeds)--Fossil binomial name paleobotanists call today's Picea breweriana, the Brewer spruce, a rather rare North America tree that now grows in natural habitat only in the Klamath Mountains of southwest Oregon and northwest California at elevations from 1,000–2,700 m (3,300–8,900 ft). DNA genetic analyses placed the Brewer spruce at the base of the Picea clade, suggesting that spruces originated in North America. This species also occurs in the middle Miocene Pyramid Formation. 0, 228, 2, 7, 169, 44, 0, 0, 0. 8) Picea lahontense (winged seeds)--A presumed extinct species of spruce whose samaras cannot be matched exactly with any living variety, although they do resemble winged seeds produced by the larger-coned spruces of eastern China. 0, 64, 0, 7, 124, 84, 0, 21, 0. 9) Pinus balfouroides (winged seeds, cones)--The paleobotanical genus-species for fossil plant material that is quite similar to the modern foxtail pine Pinus balfouriana, endemic to northern California in the Siskiyou, Scott, Salmon, Marble, Trinity, and Yolla Bolly Mountains, usually at elevations from 1,525 to 1,830 m (5000 to 6,000 feet). A disjunct subspecies of foxtail pine called Pinus austrina occurs mainly within Sequoia National Park, western slopes of the Sierra Nevada, near the headwaters of the Kings, Kern and Kaweah Rivers. 3, 0, 0, 5, 20, 67, 0, 0, 0. 10) Pinus quinifolia (winged seeds, needles)--The Miocene equivalent of the modern Western white pine Pinus monticola, which today grows in Washington state, Montana, Idaho, Nevada, Oregon, California, and Canada (Alberta-British Columbia) at elevations from 1000 m (3,280 feet) in northern regions to 1900-3000 m (6,200 to 9,842 feet) throughout its southern reaches. 0, 78, 0, 23, 111, 0, 19, 28, 0. 11) Pinus tiptoniana (needles)--Genus-species designation for paleobotanical structures comparable to the living Allepo pine Pinus halepensis, native to Morocco, Algeria, Tunisia, Spain, southern France, Malta, Italy, Croatia, Montenegro, Albania, Greece, Syria, Lebanon, southern Turkey, Jordan, Israel, and Palestine at elevations from sea level to 1,700 m (5,600 feet). 9, 0, 0, 0, 0, 0, 0, 0, 0. 12) Pseudotsuga sonomensis (winged seeds, needles)--Paleobotanical counterpart for the living Douglas-fir, Pseudotsuga menziesii, a member of the pine family (though it's not a true fir, hemlock or spruce), whose natural habitat includes British Columbia, Canada, southward through Washington, Oregon, California, and Nevada at elevations from 550m -2,900m (1,800 to 9,500 ft), although river bottom populations can live at 800 to 900 feet. A species also found in the middle Miocene Pyramid Formation. 0, 1, 2, 11, 14, 18, 0, 6, 0. 13) Tsuga mertensioides (foliage)--Binomial name applied to fossil preservations that closely resemble structures produced by the extant mountain hemlock whose current natural range extends from southern Alaska, south to western Canada, western Washington, western Oregon, northern and eastern California (Klamath and Sierra Nevada Mountains), northwestern Idaho and northwestern Montana from sea level to 1,000 m (3,300 feet) in Alaska to 2,500 to 3,050 m (8,200-10,010 ft) in California's Sierra Nevada. 0, 3, 0, 0, 0, 1, 0, 0, 0. 14) Glyptostrobus oregonensis (foliage and cones)--The paleobotanical analog to today's Chines swamp cypress Glyptostrobus pensilis, native to southeastern China, northern Vietnam, and Laos, typically preferring river banks, ponds and swamps; grows in waters up to 60 cm (24 inches) deep. 19, 0, 0, 0, 0, 0, 0, 0, 0. 15) Sequoiadendron chaneyii (twigs, foliage)--The fossil analog of the living giant sequoia/Big Tree/Sierra redwood Sequoiadendron giganteum, now confined in native habitat to several isolated groves along the moist western slopes of California's Sierra Nevada, where it prefers elevations from 900-2700 m (2,952 to 8,852 feet). 110, 0, 14, 0, 45, 176, 52, 0, 5. 16) Juniperus desatoyana (foliage)--Fossil genus-species for paleobotanical material that closely resembles plant structures produced by the modern western juniper Juniperus occidentalis, which in natural habitat extends from southeastern Washington to eastern and central Oregon, southwest Idaho, northeastern California and northwest Nevada, usually at altitudes from 800–3,000 meters (2,600–9,800 feet). 0, 0 , 0, 0, 0, 0, 0, 1, 0. 17) Acer columbianum (synonymous with A. trainii) (leaves and samaras)--Miocene genus-species for the extant Rocky Mountain maple Acer glabrum var. glabrum that in natural range occurs in Arizona, Colorado, Idaho, Montana, Nebraska, New Mexico, Nevada, South Dakota, Utah, and Wyoming--often in association with ponderosa pine, Douglas-fir, and quaking aspen. 0, 0, 0, 0, 1, 0, 0, 0, 0. 18) Acer middlegateii (synonymos with A. chaneyii) (samaras)--The Miocene variety of today's sugar maple Acer saccharum that in natural habitat can be found in southeastern Canada, southward to Minnesota, eastern Iowa, Missouri, eastern Kansas, southeastern North Dakota, northeastern South Dakota, Wisconsin, Illinois, Michigan, Indiana, Ohio, Pennsylvania, New York, Vermont, New Hampshire, Maine, Connecticut, Massachusetts, Rhode Island, northern New Jersey, northern Maryland, western Virginia, West Virginia, Kentucky, Tennessee, western North Carolina (scattered occurrences in the central part of the state), Georgia (two isolated areas in the north), and South Carolina (one disjunct area in the northwestern portion of the state). This species also occurs in the middle Miocene Pyramid Formation. 1, 0, 0, 0, 0, 0, 0, 0, 0. 19) Acer negundoides (leaves)--The binomial paleobonical name given to the modern box elder Acer negundo, one of the most widespread species of tree in America, occurring naturally in every US state, except for Washington, Oregon, Maine, and Rhode Island. It's also present in south-central Canada and ranges as far south as Guatemala. Acer negundoides also occurs in the middle Miocene Pyramid Formation. 1, 0, 0, 0, 0, 0, 0, 0, 0. 20) Acer scottiae (leaves)--Fossil genus-species for paleobotanical structures that match closely those produced by today's yellow-paint maple Acer pictum, endemic to most of China, in addition to Japan, Mongolia, and eastern Russia. 0, 0, 3, 0, 2, 0, 0, 0, 0. 21) Mahonia reticulata (leaves)--The Miocene binomial name for today's Island barberry Mahonia pinnata subsp. insularis, now found growing in the wild only on Santa Cruz Island off the coast of southern California. 1, 6, 1, 0, 0, 5, 0, 0, 0. 22) Mahonia macginitiei (leaves)--Genus-species used to describe the fossil form of the extant Tall Oregon grape Mahonia aquifolium, whose natural habitat includes southern Canada south to Washington, western Oregon, Idaho, western Montana, California, and the US Great Lakes region. 0, 4, 1, 0, 2, 2, 0, 1, 0. 23) Betula thor (leaves)--The fossil counterpart to the living paper birch (also called American white birch) Betula papyrifera, presently native to most of Canada and Alaska and portions of Washington state, Oregon, Idaho, Montana, North Dakota, South Dakota, Nebraska, Colorado (southernmost occurrence), Minnesota, Wisconsin, Michigan, Illinois, Iowa, Indiana, Ohio, New York, West Virginia, North Carolina, Pennsylvania, Connecticut, Massachusetts, and Rhode Island. It also grows throughout all of Vermont, New Hampshire, Maine, and Massachusetts. A species that is also found in the middle Miocene Pyramid Formation. 9, 0, 0, 0, 0, 0, 0, 0, 0. 24) Betula smithiana (leaves)--Paleobotanical genus-species for fossil remains of structures that are quite similar to the modern red birch Betula occidentalis, native to much of southern Canada and portions of Washington, Oregon, California (mainly northwestern areas and Sierra Nevada), Nevada (isolated populations in central and eastern sectors), northern Arizona, northern New Mexico, Utah, Colorado, Wyoming, Idaho, Montana, North Dakota, South Dakota, and western Nebraska. 0, 30, 0, 3, 6, 0, 0, 4, 0. 25) Betula vera (leaves)--The Miocene equivalent to the modern yellow birch (also called golden birch or swamp birch) Betula lutea (synonymous with Betula alleghaniensis). Likes cooler environments, typically in swamps, stream banks, deep woods, and north facing slopes from southeastern Canada southward to northeastern Minnesota, Wisconsin, northeastern Iowa, northernmost Illinois, northern Indiana, northeastern Ohio, Michigan, Pennsylvania, New York, Vermont, New Hampshire, Maine, Connecticut, Massachusetts, Rhode Island, northern New Jersey, northern Maryland, West Virginia, western half of Virginia, Kentucky, Tennessee, western North Carolina, northwestern South Carolina, and northeastern Georgia. This species also occurs in the middle Miocene Pyramid Formation. 0, 1, 0, 0, 0, 0, 0, 0, 0. 26) Betula ashleyii (leaves)--Bionmial name that paleobotanists give to the living water birch (also called red birch) Betula fontinalis (synonymous with Betula occidentalis), which in natural habitat ranges southward through southern Canada to form scattered occurrences in Washington, Oregon, California, Idaho, Nevada, Arizona, New Mexico, Colorado, Montana, Wyoming, North Dakota, South Dakota, and western Nebraska. 0, 0, 0, 0, 0, 0, 1, 0, 1. 27) Arbutus matthesii (leaves)--The paleobotanical analog to today's Pacific madrone Arbutus menziesii, native primarily to southern Canada south to California--though they are rare south of Santa Barbara County; an isolated population inhabits the western foothills of the Sierra Nevada. Arbutus matthesii also occurs in the middle Miocene Pyramid Formation. 1, 0, 0, 0, 0, 3, 2, 0, 0. 28) Arbutus idahoensis (leaves)--According to paleobotanist Daniel I. Axelrod, this is an extinct species of madrone whose closest modern-day affinity is possibly with the Texas madrone Arbutus xalapensis, native to Central America, Mexico, and the southwestern United States (Texas and New Mexico). Arbutus idahoensis is also found the middle Miocene Pyramid Formation. 0, 0, 0, 3, 3, 0, 0, 0, 0. 29) Castanopsis sonomensis (synonymous with Chrysolepis sonomensis) (leaves)--The Miocene name for the living giant chinquapin Chrysolepis chrysophylla, presently native to southern Washington southward through western Oregon to west-central California, favoring climax forests dominated by redwood, white fir, western hemlock, and Shasta red fir. Castanopsis sonomensis has also been collected from the middle Miocene Pyramid Formation. 0, 0, 1, 0, 4, 0, 0, 0, 0. 30) Lithocarpus klamathenis (synonym of Lithocarpus nevadensis) (leaves)--The fossil equivalent to the modern tanbark-oak Notholithocarpus densiflorus, endemic to California and southern Oregon. Despite its common name and the fact that it does produce acorns, it is nevertheless not a true oak. This genus-species is also found in the middle Miocene Pyramid Formation. 0, 0, 11, 0, 0, 1, 0, 1, 1. 31) Quercus pollardiana (leaves, acorns)--A presumed extinct species of oak whose fossil leaves and acorns closely resemble today's canyon live oak Quercus chrysolepis, a common component of mountainous botanical communities in California (Sierra Nevada, Coast Ranges, Klamath Mountains, and San Gabriel Mountains, for example); also occurs in southwestern Oregon, western Nevada, Arizona, southwestern New Mexico, Baja California, and Chihuahua Mexico. Has been found growing at elevations from 500 to 2,700 m (1,600 to 8,852 feet). 11, 0, 275, 0, 3, 1, 440, 0, 4. 32) Quercus simulata (leaves)--An extinct species whose fossil leaves and acorns compare with no known modern oak. Not a few paleobotanists suggest that it's possibly allied with the living willow oak Quercus phellos, native to the southeastern United States, or even the silver oak Quercus hypoleucoides, currently found in southeastern Arizona, southwestern New Mexico, and northern Mexico. 686, 0, 0, 0, 0, 0, 30, 0, 0. 33) Crataegus gracilens (leaves)--Paleobotanical genus-species applied to the fossil variety of today's scarlet hawthorn Crataegus coccinea, native to most of eastern Canada, south to Connecticut, Illinois, Indiana, Maine, Massachusetts, Maryland., Michigan, Minnesota, North Carolina, New Hampshire, New York, Ohio, Pennsylvania, Rhode Island, Virginia, Vermont, Virginia, and Wisconsin. 0, 0, 0, 0, 0, 2, 0, 0, 0. 34) Gymnocladus dayana (leaves)--The fossil counterpart to the living Kentucky coffeetree Gymnocladus dioicus, a rather rare tree that is native to northern Kentucky, central and western Tennessee, extreme northwest Alabama, Arkansas, Missouri, Oklahoma, eastern Kansas, Iowa, southern Minnesota, southeastern Wisconsin, southern Michigan, Indiana, western to southern Ohio, western Pennsylvania, central New York, scattered areas in West Virginia, one disjunct place in eastern Nebraska, one isolated spot in western North Carolina near the West Virginia border, and Ontario Canada. Typically favors floodplains and river valleys, though it's known to populate rocky hillsides and limestone woods, as well. 0, 1, 0, 0, 0, 0, 0, 0, 0. 35) Platanus bendirei (leaves)--This is what paleobotanists call fossil specimens that match today's California sycamore Platanus racemosa (also referred to as the western sycamore), a chiefly riparian specimen that ranges from western and eastern California--northernmost occurrence is in Humbolt County, with scattered patches in the Sierra Nevada--south to northern Baja California Mexico. This species also occurs in the middle Miocene Pyramid Formation. 1, 0, 0, 0, 0, 0, 0, 0, 0. 36) Zelkova brownii (leaves)--Genus-species for the paleobotanical analag of the living Japanese zelkova Zelkova serrata, native to Japan, Korea, Taiwan, and Manchuria; favors riparian habitats and rich soils at the base of mountains. 0, 1, 0, 0, 0, 0, 0, 0, 0. 37) Amalanchier nevadensis (leaves)--The scientific binomial name for fossil plants that show great similarity to today's western serviceberry (pigeon berry) Amelanchier alnifolia, which grows naturally in central and southern Alaska, western to central Canada, Washington, Oregon, northwestern California, Nevada (scattered occurrences), Idaho, Montana, North Dakota, South Dakota, northern Nebraska, Wyoming, Colorado, and Utah from sea level (north) to 2,600 m (8,530 feet) in California and 3,400 m (11,200 feet) in the Rocky Mountains. 0, 40, 0, 0, 19, 6, 0, 1, 0. 38) Ceanothus leitchii (leaves)--Fossil binomial name for the modern Tobacco brush (snowbrush ceanothus) Ceanothus velutinus, widespread in Alberta and British Columbia, south through Washington, Oregon, California, Idaho, Montana, Wyoming, South Dakota, Utah, and Colorado from sea level to elevations from 450 to 3,050 meters (1,476 to 10,000 feet). 0, 2, 0, 0, 5, 6, 0, 0, 0. 39) Rhamnus columbiana (leaves) This is the paleobotanical name given to fossils of the living California coffeeberry (a buckthorn) Frangula californica (used to be called Rhamnus californica); presently native to California, southwestern Oregon, southern Nevada, Arizona, western New Mexico, and Baja California Mexico, preferring oak woodlands and chaparral habitats. 1, 0, 0, 1, 0, 0, 0, 0, 0. 40) Amelanchier alvordensis (leaves)--The Miocene designation for the modern western serviceberry Amelanchier alnifolia, whose native habitat ranges from Alaska through western to central Canada, and then south to Washington, Oregon, northwestern California, Idaho, mainly western and eastern Montana, North Dakota, South Dakota, western Minnesota, northwestern Iowa, northern Nebraska, Wyoming, Colorado, Utah, and scattered patches in Nevada at elevations from sea level in its northernmost ranges to 2,600 m (8,530 feet) in California) and 3,400 m (11,200 feet) in the Rocky Mountains. 0, 40, 0, 7, 19, 0, 0, 1, 0. 41) Cercocarpus antiquus (synonymous with Cercocarpus nevadensis) (leaves)--The binomial term paleobotanists give to the fossil variety of today's mountain mahogany Cercocarpus betuloides, distributed natively in California, Oregon, Arizona, northwestern New Mexico, and Baja California Mexico, where it typically prefers chaparral communities in summer dry foothills and mountains. 2, 43, 2, 36, 172, 0, 1, 7, 0. 42) Cercocarpus linearifolius (leaves)--The fossil representative of the living curl-leaf mountain mahogany Cercocarpus ledifolius, now native to extreme southeastern Washington, Oregon, California (mainly western Sierra Nevada foothills), Nevada, northern Arizona, Utah, western Colorado, western Montana, and northern Baja California Mexico, where it typically forms scattered groves in pinyon pine-juniper-sagebrush ecosystems at elevations from 600 to 3,000 m (2,000 to 9,800 feet), preferring well-drained sandy or grainy soils in areas that receive on average 15–26 cm (6 to 10 inches) precipitation per year. 0, 2, 0, 0, 0, 1, 0, 0, 0. 43) Heteromeles sonomensis (synonymous with H. stenophylla) (leaves)--The paleobotanical name given to fossil specimens of the extant toyon (also called Christmas berry) Heteromeles arbutifolia, native to coastal sagebrush and chaparral communities of southwestern Oregon southward to California's Coast Ranges (beginning in Humbolt County) and Transverse Ranges; a disjunct population inhabits the western foothills of the Sierra Nevada. Its natural range extends to northern Baja California Mexico. 0, 0, 0, 1, 3, 6, 0, 0, 0. 44) Holodiscus idahoensis (leaves)--Paleobotanical binomial name for fossil specimens of the living oceanspray Holodiscus discolor var. glabrescens, which in native habitat extends from western Canada south to Washington, Oregon, California, Nevada, Idaho, Montana, Utah, and Colorado; prefers old-growth conifer, seral and old-growth hardwood, mixed-riparian, and mixed-shrub environments. 0, 0, 0, 1, 13, 0, 0, 2, 0. 45) Cornus ovalis (leaves)--The genus-species provided for fossils that match structures produced by the modern California dogwood/western dogwood Cornus californica (synonymous with Cornus sericea ssp. occidentalis), endemic to California--primarily, the central to northern Sierra Nevada and the northwestern to southern coastal areas at elevations from sea level to 2,438 meters (8,000 feet). 0, 1, 0, 0, 0, 0, 0, 0, 0. 46) Prunus chaneyii (leaves)--This is the fossil equivalent to the extant Western chokecherry Prunus virginiana var. demissa, presently native to northern, southern and central California, preferring in natural habitat rocky slopes at elevations from 0-8200 feet. Prunus chaneyii also occurs in the middle Miocene Pyramid Formation. 1, 3, 0, 1 0, 1, 0, 1, 0. 47) Robinia californica (leaves)--The paleobotanical analog to the living New Mexican black locust Robinia neomexicana, endemic to California (eastern Mojave Desert and San Gabriel Mountains sky island pinyon pine-juniper habitats, usually at elevations higher than 1,524 meters--5,000 feet), southeastern Nevada, southwestern Utah, Arizona, New Mexico, central and southeastern Colorado, New Mexico, extreme western Texas, and northern Mexico at altitudes between 1,200 and 2,600 meters (4,000-8,500 feet) along streams, at the bottoms of valleys, and the sides of canyons. 0, 2, 0, 1, 1, 2, 0, 0, 0. 48) Rosa shornii (leaves)--Named in honor of paleobotanist Howard Schorn, former collections manager of fossil plants in the University California Museum of Paleontology, Berkeley, California; the Miocene analog to the modern California wildrose Rosa californica now native to California (notably, coast and foothills up to elevations of 6,000 feet), Oregon and northern Baja California Mexico. 0, 1, 0, 0, 0, 0, 0, 0, 0. 49) Sorbus cassiana (leaves)--The genus-species designation for fossil plant structures quite similar to those generated by the living whitebeam (mountain ash) Sorbus pohuashanensis (synonymous with Sorbus aucuparia subsp. pohuashanensis), presently endemic to north and northeast China. 0, 0, 0, 1, 4, 1, 0, 0, 0. 50) Vaccinium sophoroides (leaves)--Paleobotanical name for the extant Vaccinium corymbosum, the northern highbush blueberry, native to Connecticut, Maine, Massachusetts, New Hampshire, Rhode Island, Vermont, New York, Pennsylvania, Maryland, New Jersey, West Virginia, Virginia, North Carolina, South Carolina, Georgia, Florida, Tennessee, Kentucky, Mississippi, Alabama, Louisiana, Texas, Oklahoma, Kansas, Missouri, Iowa, Wisconsin, Michigan, Illinois, Indiana, Ohio, and Canada. 1, 0, 0, 0, 0, 1, 0, 0, 0. 51) Leucothoe nevadensis (leaves)--The fossil genus-species for today's Sierra laurel Leucothoe davisiae, endemic to extreme southwest Oregon and the Sierra Nevada and Klamath Mountains of California, inhabiting mountain bogs and wetlands at altitudes from 400--3,000 meters (1,312 to 9,842 feet). 0, 0, 0, 0, 0, 1, 0, 0, 0. 52) Lyonothamnus parvifolia (synonymous with L. cedrusensis) (leaves)--A presumably extinct species whose fossil leaves mostly match those produced by the extant Santa Cruz Island Ironwood Lyonothamnus floribundus ssp. asplenifolius, presently endemic to Santa Cruz Island off the coast of southern California, where it likes chaparral and oak woodlands in rocky coastal canyons at elevations from 100 to 1,600 feet (30 to 487 meters). 2, 3, 0, 0, 0, 0, 2, 5, 2. 53) Populus cedrusensis (leaves)--Paleobotanical designation for fossil material that closely resembles the modern guerigo (also called white bark cottonwood) Populus brandegeei (synonymous with Populus monticola), known in native range from nouthern Baja California to east-central Sonora Mexico. 0, 10, 0, 1, 1, 0, 0, 0. 54) Populus eotremuloides (synonymous with P. bonhamii) (leaves, one capsule)--The scientific genus-species designation for paleobotanical remains that closely resemble the modern black cottonwood Populus balsamifera. subsp trichocarpa. which today populates portions of southern Alaska southward through western Canada, Washington, Oregon, California (northwestern, western foothills of the Sierra Nevada, and spotty places in southern areas), northern to central Idaho, western Montana, and northern Baja California Mexico; also documented in scattered patches in southeastern Alberta, eastern Montana, western North Dakota, western Wyoming, Utah, and Nevada. 5, 156, 4, 38, 66, 42, 14, 53, 0. 55) Populus payettensis (leaves)--The Miocene analog to the modern narrowleaf cottonwood Populus angustifolia, which today has a wide geographic range of natural habitat--southern Canada, southward to Montana, Idaho, Wyoming, Colorado, Utah, Arizona, New Mexico, northeast Nevada, eastern California, southeastern Oregon, western Texas, and northern Mexico--mainly along streams and creeks at elevations from 3,900 to 7,900 feet (1,200 to 2,400 m). 0, 2, 0, 0, 0, 6, 0, 0, 3. 56) Populus pliotremuloides (leaves)--Paleobotanical genus-species for fossil leaves that match the living quaking aspen Populus tremuloides, one of the most widespread trees in North America, ranging in native populations from Alaska through most of Canada, south to every US state except Kansas, Oklahoma, Arkansas, Louisiana, Alabama, Mississippi, Florida, Georgia, South Carolina, North Carolina, Tennessee, and Kentucky. 0, 10, 0, 0, 0, 2, 0, 1, 1. 57) Salix churchillensis (leaves)--The Miocene paleobotanical designation for the modern sandbar willow (coyote willow) Salix exigua subspecies exigua, which now populates places southward from southern Canada to eastern Washington, eastern Oregon, eastern California, Idaho, Nevada, southwestern Montana, western Wyoming, Utah, Colorado, Arizona, New Mexico, western Texas, and northern Mexico. Salix churchillensis also occurs in the middle Miocene Pyramid Formation. 11, 17, 0, 1, 10, 9, 0, 2, 2. 58) Salix desatoyana (leaves)--Genus-species paleoboanists give to fossil leaves that show little variation from living foliage of the black willow Salix nigra, native to southeastern Canada, southern Maine, New Hampshire, Vermont, Massachusetts, Connecticut, Rhode Island, New Jersey, Maryland, North Carolina, South Carolina, Georgia, northwest Florida, Mississippi, Alabama, Louisiana, eastern Texas, Oklahoma, Arkansas, Tennessee, Kentucky, Pennsylvania, New York, southern Michigan, Ohio, Indiana, Illinois, Iowa, Wisconsin, Missouri, southeastern Minnesota, eastern Kansas, central to southern California, Arizona, New Mexico, southern Utah, and northern Mexico. 0, 16, 0, 2, 6, 2, 0, 1, 0. 59) Salix inquirenda (leaves)--The Miocene equivalent of today's Salix lasiandra, the Pacific willow, widely distributed from Alaska southward through western to central Canada, Washington, Oregon, California, Idaho, Montana, Wyoming, Colorado, Arizona, and New Mexico. A species that also occurs in the middle Miocene Pyramid Formation. 0, 2, 0, 2, 0, 0, 0, 0, 0. 60) Salix laevigitoides (leaves)--The fossil equivalent to the modern red willow Salix laevigata, which today is found mainly from Cape Mendocino (California coast), southward into northern Baja California Mexico. It also occurs in the lower elevation western foothills of California's Sierra Nevada, southern Nevada, southwestern Utah, and central Arizona. 0, 15, 0, 10, 12, 18, 0, 80, 2. 61) Salix pelviga (leaves)--This is the fossil genus-species for the living dusky willow Salix melanopsis, native to British Columbia and Alberta southward to California and Colorado, where it likes moist riverbanks and subalpine mountain meadows. This species is also found in the middle Miocene Pyramid Formation. 1, 8, 1, 23, 20, 68, 1, 0, 3. 62) Salix storeyana (synonymous with S. knowltonii) (leaves)--Paleobotanical designation for the extant Lemmon's willow Salix lemmonii, native to southeastern Washington, eastern two-thirds of Oregon, northern Nevada, Idaho, southwestern Montana, northwestern Wyoming, and eastern California (with an isolated population in inland southern California). 0, 12, 3, 1, 0, 0. 0, 0, 0. 63) Salix wildcatensis (leaves)--Scientific bionmial name for leaves that show no appreciable difference from today's arroyo willow Salix lasiolepis, which in native habitat currently populates portions of central to southern Washington, Oregon, western Idaho, California, Nevada, central to southwestern Utah, Arizona, southern New Mexico, western Texas, and north Mexico, typically in riparian zones situated in canyons and valleys, pond shores, marshes, and wetlands. 0, 0, 0, 0, 0, 3, 1, 0, 0. 64) Alnus spokanensis (leaves, cones): The fossil equivalent of the modern speckled alder Alnus incana subspecies rugosa, which today ranges in natural habit from Canada to northern North Dakota, Minnesota, Wisconsin, Michigan, northeastern Iowa, northern Illinois, northern Indiana, northern Ohio, Pennsylvania, western Maryland, eastern West Virginia, New York, Connecticut, Rhode Island, Massachusetts, Vermont, New Hampshire, and Maine. This species also occurs in the middle Miocene Pyramid Formation. 0, 1, 0, 0, 0, 0, 3, 0, 0. 65) Garrya idahoensis (leaves): Fossil name given to plant structures that most resemble the living California fever bush Garrya fremontii, which inhabits California, western Oregon, and southern Washington in both forested and chaparral associations. 0, 0, 0, 0, 1, 0, 0, 0, 0. 66) Equisetum sp. (stems)--Structures that closely resemble stems from the living horsetail, genus Equisetum, probably allied with the scouring rush of worldwide geographic botanical distribution. 1, 0, 0, 0, 0, 0, 0, 0, 0. 67) Typha lesquereuxii (leaves)--The genus-species paleobotanists give to fossil structures that match those produced by the extant bulrush (broadleaf cattail) Typha latifolia, native to every US state except Hawaii, Canada, North and South America, Europe, Eurasia, and Africa. It is an obligate wetlands botanic specimen, requiring watery environments to propagate. 4, 0, 1, 0, 0, 0, 0, 0, 0. 68) Pteridium calabazensis (pinnules)--The paleobonical designation for fossil material that closely resembles the living bracken (eagle fern) Pteridium aquilinum, presently found on all continents except Antarctica. 0, 2, 0, 0, 0, 0, 0, 0, 0. I arrived back home reinvigorated with paleobotanical zeal after my excellent expedition. Consecutive days of solitary isolation amidst ancient strata that told true tales of abundant flourishing vegetation in a land left with arid botanic austerity had inspired a consuming need to learn all that I could of what once existed there all those many millions of years ago, to move through space and time in a quantum-like delirium of amazed revelry, seeing now the infinity of geologic time transpire in a continuum of imagination's revelatory reality, where fir and spruce and maple and birch and giant sequoias rise once again above a cool clear lake in the desert as a distant volcano only awaits its moment to preserve eternity. |
 |
 |
| Click on the images for larger pictures. Left and right: Two views of the fossil locality in the middle Miocene Pyramid Formation not far from Reno, Great Basin Desert, Nevada, a site that yields to paleobotanical investigations some 41 species of 15.6 million year-old plants, preserved in bone-white diatomites (composed almost entirely of diatoms, a microscopic single celled photosynthesizing variety of algae). Note 1979 American Motors CJ7 jeep for scale. That trench provides comfortable and convenient access to the plant-yielding sedimentary rocks. Dramatic peak in background is a quartz dacite plug, solidified in the vent of a volcano that formed here 21 million years ago. |
 |
 |
| Click on the images for larger pictures. Left and right--An enthusiast of paleobotany explores the middle Miocene Pyramid Formation in the vicinity of Reno, Great Basin Desert, Nevada, where diatomites (a rock type composed almost entirely of diatoms, a photosynthesizing microscopic single celled kind of algae) deposited approximately 15.6 million years ago produce 41 species of middle Miocene plants. |
 |
 |
| Click on the images for larger pictures. Left--Viewing perspective from a typical surface outcropping of highly eroded diatomite (composed almost entirely of diatoms, a microscopic photosynthesizing single-celled variety of algae) in the middle Miocene Pyramid Formation, as exposed in the neighborhood of Reno, Nevada, Great Basin Desert, where folks interested in paleobotanical adventure can find 41 species of fossil plants. Hills in distance are composed of the 22 to 25 million year old volcanic Tule Peak rhyolite. Right--Turnoff at left leads to the middle Miocene Pyramid Formation locality not far from Reno, Nevada, Great Basin Desert, where some 41 species of 15.6 million year old plants have been identified from leaf, seed, and conifer foliage impressions. A Google Street Car perspecitve that I edited and processed through photoshop. |
 |
 |
| Click on the images for larger pictures. Left--A paleobotany aficionado investigates a 13.3 to 14.8 million year old fossil locality in the Chloropagus Formation, in the vicinity of Reno, Nevada, Great Basin Desert, which yields a rather rich fossil flora comprised of 68 species of middle Miocene plants. Right--A view roughly northward to hills (foreground) underlain by the fossil plant-bearing middle Miocene Chloropagus Formation, dated at 13.3 to 14.8 million years old, which provides paleobotany adventurers with a wealth of leaves, seeds, needles, and fascicles from 68 species of plants. The locality lies in the neighborhood of Reno, Nevada, The Biggest Little City In The World, Great Basin Desert, Nevada. Note light dusting of snow on mountain tops in distance. |
 |
 |
| Click on the images for larger pictures. Left--A classic fossil plant locality in the middle Miocene Chloropagus Formation (yields 68 species of ancient plants), situated in the neighborhood of Reno, Great Basin Desert, Nevada, a specific site that records four individual superposed fossil floras, each horizon deposited one atop the other in an unfaulted, normal stratigraphic succession, providing paleobotanists with a wonderful opportunity to analyze the evolution of plant communities through a specific period of geologic time. Note red backpack for scale. Right--A geology rock hammer provides scale for an exposure of fossil plant-bearing lacustrine shales in the middle Miocene Chloropagus Formation, deposited some 13.3 to 14.8 million years ago in the vicinity of present-day Reno, Nevada, in localized shallow lakes and ponds that were periodically subjected to extrusive volcanic activity. |
 |
 |
| Click on the images for larger pictures. Left and right--Two views of the same locality, where a paleobotanically motivated person searches for fossil plants in the middle Miocene Chloropagus Formation, in the vicinity of Reno, Nevada, Great Basin Desert. 68 species of 13.3 to 14.8 million year old plants have been identified here from leaves, seeds, needles, fascicles, cones, cone scales, and foliage. Note snow on hills in distance. |
 |
 |
| Click on the images for larger pictures. Left--Typical exposure of fossiliferous shales in the middle Miocene Chloropagus Formation, not far from Reno, Nevada, Great Basin Desert, a geologic rock unit that yields leaves, seeds, needles, fascicles, foliage, cones and cone scales from 68 species of 13.3 to 14.8 million year old plants. Right--Dirt road at left delivers paleobotatany aficionados to fossil-bearing outcrops of the 13.3 to 14.8 million year old Chloropagus Formation in the neighborhood of Reno, Nevada, Great Basin Desert, where the leaves, seeds, needles, and foliage from 68 species of fossil plants have been recovered. A Google Street Car perspecitive that I edited and processed through photoshop. |
 |
 |
|
Click on the images for larger pictures. Left--View is eastward up the Yuba River, western foothills of California's Sierra Nevada, to a vegetational ecotone--a transitional botanic association situated between a mixed conifer forest and a broadleaf sclerophyll community (chaparral)--where several varieties closely allied to fossil types found in the 15.6 million year old middle Miocene Pyramid Formation now grow, among them--Ponderosa pine (Pinus ponderosa), Douglas-fir (Pseudotsuga menziesii), incense cedar (Calocedrus masonii), Pacific madrone (Arbutus menziesii), chinquapin (Chyrsolepis), tanbark oak (Lithocarpus), Western sycamore (Platanus racemosa), box elder (Acer negundo), bigleaf maple (Acer macrophyllum), Oregon ash (Fraxinus oregona) dusky willow (Salix melanopsis), sandbar willow (Salix exigua), and wild rose (Rosa). A Google Street Car perspective that I edited and processed through photoshop. Right--Near Metcalf Bottoms, southeast of Townsend, Tennessee, a region that supports a rich deciduous hardwood forest with many plant species similar to varieties recovered from the middle Miocene Pyramid Formation, 15.6 million years old, including species of maple (Acer), buckeye (Aesculus), alder (Alnus), birch (Betula), Kentucky yellow tree (Cladastris), ash (Fraxinus), American hophornbean (Ostrya), and chokecherry (Prunus). A Google Street Car perspective that I edited and processed through photoshop. |
 |
 |
|
Click on the images for larger pictures. Left--A scene at Calaveras Big Trees State Park (North Grove area), western foothills of California's Sierra Nevada, amidst a rich mixed conifer forest--with a giant sequoia at right--a place that supports several conifers, sclerophylls, and deciduous kinds closely allied with fossil types found in the 13.3 to 14.8 million year old Chloropagus Formation in the vicinity of Reno, Nevada, including: giant sequoia (Sequoiadendron), fir (Abies), pine (Pinus), Douglar-fir (Pseudotsuga), madrone (Arbutus), tanbark oak (Lithocarpus), canyon live oak (Quercus), serviceberry (Amalanchier), buckbrush (Ceanothus), black laurel (Leucothoe), Oregon grape (Mahonia), western chokecherry (Prunus), wild rose (Rosa), and willow (Salix). A public domain photograph courtesy John J. O'Brien; I edited and processed it through photoshop. Right--Sugar Lake, Salmon Mountains--a branch of the Klamath Mountains--northwestern California, amidst the southern sector of the Miracle Mile, a world famous square mile of gorgeous geography that supports one of the most diverse conifer assemblages on Earth, a list that includes foxtail pine (Pinus balfouriana), whitebark pine (Pinus balfouriana), western white pine (Pinus monticola), Jeffrey pine (Pinus jeffreyi), ponderosa pine (Pinus ponderosa), lodgepole pine (Pinus contorta), sugar pine (Pinus lambertiana), white fir (Abies concolor), Shasta fir (Abies magnifica), subalpine fir (Abies lasiocarpa), Engelmann spruce (Picea engelmannii), Brewer spruce (Picea breweriana), mountain hemlock (Tsuga mertensiana), Douglas-fir (Pseudotsuga menziesii), Pacific yew (Taxus brevifolia), incense-cedar (Calocedrus decurrens), common juniper (( communis), and western juniper (Juniperus occidentalis). All told, some 32 species of conifers have been identified from the Klamath Mountains region. A Google Earth image--photographer unknown-- that I edited and processed through photoshop. |
 |
 |
|
Click on the images for larger pictures. Left--A view roughly northwest along the eastern slopes of Cone Peak, Santa Lucia Range, California, an area that supports several plant species similar to types that occur in the 13.3 to 14.8 million year old Chloropagus Formation in the neighborhood of Reno, Nevada, Great Basin Desert, including Shasta red fir (Abies magnifica), sugar pine (Pinus lambertiana), madrone (Arbutus), canyon live oak (Quercus), tanbark oak (Lithocarpus), Oregon grape (Mahonia), mountain mahogany (Cercocarpus), dogwood (Cornus), toyon (Heteromoles), oceanspray (Holodiscus), pigeon berry (Amalanchier), western chokecherry (Prunus), wild rose (Rosa), and buckthorn (Rhamnus). A Google Earth photograph courtesy Ben Mayne; I edited and processed it through photoshop. Right--A scene in northern Idaho, where many conifers and shrubs allied with fossil kinds found in the 13.3 to 14.8 million year old Chloropagus Formation now grow, including larch (Larix), mountain hemlock (Tsuga), fir (Abies), Douglas-fir (Pseudotsuga), western serviceberry (Amalanchier), scarlet hawthorn (Crataegus), oceanspray (Holodiscus), oregon grape (Mahonia), wild rose (Rosa), willow (Salix), western chokecherry (Prunus), and whitebeam (Sorbus). A Google Street Car perspective that I edited and processed through photoshop. |
 |
 |
 |
|
Click on the images for larger pictures. Left and far right--Fossil leaves of an extinct oak Quercus simulata preserved on diatomite (an often economically lucrative rock type composed primarily of diatoms, a microscopic single celled photosynthesizing variety of algae) from the middle Miocene Pyramid Formation, collected by the author from a locality not far from Reno, Nevada, Great Basin Desert. Its leaves and occasional acorn cups match no known living oak, though paleobotanist Daniel I. Axelrod, who collected loads of Q. simulata over the course of several decades, postulated that because it occurs almost exclusively in lacustrine deposits, perhaps its closest living counterpart is the extant willow oak Quercus phellos, native to the southeastern United States. Others have pointed out that it also resembles the modern silver oak Quercus hypoleucoides, native to southeastern Arizona, southwestern New Mexico, and northern Mexico. Middle--A carbonized twig that belongs to what paleobotanist Daniel I. Axelrod called Chamaecyparis cordillerae (synonym of C. sierrae), the Miocene equivalent of today's Lawson cypress, Chamaecyparis lawsiana, now native to southwestern Oregon and the Klamath Mountains of northwest California at elevations from sea level to 4,900 feet (1,500 m); collected by the author from the 13.3 to 14.8 million year old middle Miocene Chloropagus Formation in the neighborhood of Reno, Nevada, Great Basin Desert. |
 |
 |
 |
|
Click on the images for larger pictures. Left and middle--Leaves from the extinct oak Quercus simulata, possibly allied with the living willow oak Quercus phellos, native to the southeastern United States, or even the silver oak Quercus hypoleucoides, presently found in southeastern Arizona, southwestern New Mexico, and northern Mexico, collected by the author from the 13.3 to 14.8 million year old middle Miocene Chloropagus Formation not far from Reno, Nevada, Great Basin Desert. Right--A leaf that resembles what paleobotanist Daniel I. Axelrod called Salix inquirenda, the Miocene equivalent of today's Salix lasiandra, the Pacific willow, which attains a wide geographic distribution, ranging from Alaska southward through west to central Canada to the following US states: Washington, Oregon, California, Idaho, Montana, Wyoming, Colorado, Arizona, and New Mexico; collected by the author in the neighborhood of Reno, Nevada, Great Basin Desert, from the 13.3 to 14.8 million year old middle Miocene Chloropagus Formation. |
 |
 |
|
Click on the images for larger pictures. Left--A winged seed from the presumably extinct spruce Picea magna, although a few paleobotanists suggest that it does resemble the living tigertail spruce Picea torano, now native to Japan. Collected by the author from exposures of the middle Miocene Chloropagus Formation, around 13.3 to 14.8 million years old, in the vicinity of Reno, Nevada, Great Basin Desert. Right--Foliage from Sequoiadendron chaneyii--the fossil analog of the living giant sequoia/Big Tree/Sierra redwood Sequoiadendron giganteum, now confined in native habitat to several isolated groves along the moist western slopes of California's Sierra Nevada, where it prefers elevations from 900-2700 m (2,952 to 8,852 feet); from the middle Miocene Chloropagus Formation (roughly 13.3 to 14.8 million years old) near Reno, Nevada, Great Basin Desert. Photograph courtesy Themodoccypress and used under the parameters provided by the Creative Commons Attribution-Share Alike 4.0 International license. I edited and processed the photograph through photoshop. |
 |
 |
|
Click on the images for larger pictures. Left--Part and counterpart of a leaf from the extinct oak Quercus simulata, possibly allied with the living willow oak Quercus phellos, native to the southeastern United States, or even the silver oak Quercus hypoleucoides, presently found in southeastern Arizona, southwestern New Mexico, and northern Mexico, collected by the author from the 13.3 to 14.8 million year old middle Miocene Chloropagus Formation not far from Reno, Nevada, Great Basin Desert. Right--Photograph courtesy an individual who goes by the cyber-name JimTH; I edited and processed the images through photoshop. A leaf from Alnus latahensis, the Miocene counterpart to the living Alnus maritima, the seaside alder or brook alder now found naturally in Oklahoma, Georgia, Maryland, and Delaware; it is the only North American member of the genus Alnus that blooms during autumn--all other alders bloom in spring. From the 13.3 to 14.8 million year old middle Miocene Miocene Pyramid Formation, as exposed in the vicinity of Reno, Nevada, Great Basin Desert. Note that with careful paleontological prepping technique, a mostly complete fossil leaf could likely be obtained here by carefully removing the diatomite matrix layer that partially obscures the specimen. |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. Fossil leaves from the 15.6 million year old middle Miocene Pyramid Formation, from a locality not far from Reno, Nevada, Great Basin Desert. Left--Rosa shornii: Named in honor of paleobotanist Howard Schorn, former collections manager of fossil plants in the University California Museum of Paleontology, Berkeley, California; the Miocene analog to the modern California wildrose Rosa californica now native to California (notably, coast and foothills up to elevations of 6,000 feet), Oregon and northern Baja California Mexico. Middle--Quercus simulata: A presumed extinct species of oak whose leaves and occasional acorn cups match no known living species, though several paleobotanists postulate that perhaps its closest living counterpart is the extant water oak Quercus phellos, native to the southeastern United States. Others have pointed out that it also resembles the modern silver oak Quercus hypoleucoides, native to southeastern Arizona, southwestern New Mexico, and northern Mexico. Right--Alnus latahensis: This is the Miocene counterpart to the living Alnus maritima, the seaside alder or brook alder now found naturally in Oklahoma, Georgia, Maryland, and Delaware; it is the only North American member of the genus Alnus that blooms during autumn--all other alders bloom in spring. |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and process them through photoshop. Fossil leaves from the 15.6 million year old middle Miocene Pyramid Formation, secured from a specific locality in the vicinity of Reno, Nevada, Great Basin Desert. Left--Lithocarpus nevadensis: The fossil equivalent to the extant tanbark-oak Notholithocarpus densiflorus, presently native to California and southern Oregon. Despite its common name and the fact that it does produce acorns, it is nevertheless not a true oak. Middle--Chrysolepis sonomensis: The paleobotanical name for the living giant chinquapin Chrysolepis chrysophylla, endemic to southern Washington southward through western Oregon to west-central California, favoring climax forests dominated by redwood, white fir, western hemlock, and Shasta red fir. Right--Cladrastis lariverisii: The Miocene analog for the living Kentucky yellowwood Cladastris lutea (syonymous with Cladrastis kentukea), which today grows in the wild only in Kentucky, Tennessee, Arkansas, Mississippi, Indiana, western Oklahoma, Alabama, western North Carolina, western Virginia, northern Georgia, and southern Missouri. |
 |
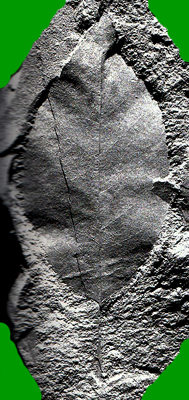 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and process them through photoshop. Fossil leaves from the 15.6 million year old middle Miocene Pyramid Formation, found at a specific locality in the vicinity of Reno, Nevada, Great Basin Desert. Left--Taxodium oregonensis: The Miocene genus-species of Taxodium distichum, the modern bald (swamp) cypress, now native to the warm, humid southeastern United States with annual precipitation levels of 760 mm or 30 inches (eastern Texas) to 1,630 mm--64 inches (Gulf Coast). Middle--Prunus chaneyii: This is the fossil equivalent to the extant Western chokecherry Prunus virginiana var. demissa, presently native to northern, southern and central California, preferring in natural habitat rocky slopes at elevations from 0-8200 feet. Right--Aesculus preglabra: Paleotanical genus-species for the living Ohio buckeye Aesculus glabra whose natural geographic distribution includes populations in Ohio, southern Michigan, eastern Pennsylvania, Tennessee, northern Alabama, Mississippi (one isolated spot near the Alabama border), central to northeastern Texas, eastern half of Oklahoma, northwestern Arkansas, western Kansas, Missouri, southern Iowa, Illinois, and Indiana. |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil leaves from the 15.6 million year old middle Miocene Pyramid Formation, found at a specific locality in the vicinity of Reno, Nevada, Great Basin Desert. Left--Populus bonhamii: The Miocene genus-species of Populus balsamifera, the balsam poplar (northernmost hardwood in North America), now native to most of Canada and the following US states: Alaska, northeastern Oregon, central and northwestern Idaho, scattered in five isolated areas in Montana, Wyoming, north to central Colorado, northwestern Nebraska, western and northeastern South Dakota, southeastern and northern North Dakota, northern half of Minnesota, northern Iowa, Wisconsin, Michigan, northeastern Illinois, northwestern Indiana, northeastern Ohio, Pennsylvania, northeastern West Virginia, western Maryland, northeastern Connecticut, New York, Vermont, northern half of New Hampshire, and Maine. Middle--Alnus spokanensis: The fossil equivalent of the modern speckled alder Alnus incana subspecies rugosa, which today ranges in natural habit from Canada southward to the following US states: northern North Dakota, Minnesota, Wisconsin, Michigan, northeastern Iowa, northern Illinois, northern Indiana, northern Ohio, Pennsylvania, western Maryland, eastern West Virginia, New York, Connecticut, Rhode Island, Massachusetts, Vermont, New Hampshire, and Maine. Right--Acer glabroides: A species analogous to the living red maple Acer rubrum, presently distributed in geographic range from southeastern Canada southward into the following US states: eastern Texas, eastern Oklahoma, Missouri, Minnesota, Michigan, Wisconsin, Indiana, southern Illinois, Ohio, Pennsylvania, New York, Connecticut, Vermont, New Hampshire, Maine, New Jersey, Maryland, Rhode Island, Massachusetts, West Virginia, Virginia, North Carolina, South Carolina, Georgia, Florida, Mississippi, Alabama, Louisiana, Arkansas, Tennessee, Kentucky. |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil leaves from the 15.6 million year old middle Miocene Pyramid Formation, found at a specific locality in the vicinity of Reno, Nevada, Great Basin Desert. Left-- Salix inquirenda: The Miocene equivalent of today's Salix lasiandra, the Pacific willow, which attains a wide geographic distribution, ranging from Alaska southward from western to central Canada through Washington, Oregon, California, Idaho, Montana, Wyoming, Colorado, Arizona, and New Mexico. Middle--Betula thor: The fossil counterpart to the living paper birch (also called American white birch) Betula papyrifera, now native to almost all of Canada and most of Alaska, southward to parts of the following US states: Washington, Oregon, Idaho, Montana, North Dakota, South Dakota, Nebraska, Colorado (southernmost occurrence), Minnesota, Wisconsin, Michigan, Illinois, Iowa, Indiana, Ohio, New York, West Virginia, North Carolina, Pennsylvania, Connecticut, Massachusetts, and Rhode Island. It also occurs throughout all of Connecticut, Vermont, New Hampshire, and Maine. Right--Fraxinus caudata: The paleobotanical genus-species for the modern Oregon ash Fraxinus oregona (synonymous with Fraxinus latifolia), native to the US west coast from western Washington southward through western Oregon to northwest and central California; it prefers moist loose soils at elevations from sea level to 900 m (3,000 feet--in the north) to 1,700 m (5,600 feet--in California). |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil plants from the 14.8 to 13.3 million year old middle Miocene Chloropagus Formation, found in the vicinity of Reno, Nevada, Great Basin Desert. Left--Winged seeds (samaras) from the presumably extinct spruce Picea magna, although a few paleobotanists suggest that it does resemble the living tigertail spruce Picea torano, now native to Japan. Middle--Fascicles (bundles of needles) from what paleobotanists call Pinus quinifolia. the Miocene analog of the modern Western white pine Pinus monticola, which today grows in the western United States--Washington, Montana, Idaho, Nevada, Oregon, California--and Canada (Alberta-British Columbia) at elevations from 1000 m (3,280 feet) in northern regions to 1900-3000 m (6,200 to 9,842 feet) throughout its southern reaches. Right--Foliage from Sequoiadendron chaneyii, the fossil equivalent of today's giant sequoia/Big Tree/Sierra redwood Sequoiadendron giganteum, now confined in native habitat to several isolated groves along the moist western slopes of California's Sierra Nevada, primarily at elevations from 900-2700 m (2,952 to 8,852 feet). |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil plants from the 14.8 to 13.3 million year old middle Miocene Chloropagus Formation, found in the neighborhood of Reno, Nevada, Great Basin Desert. Left--Fascicles (bundles of needles--at top) and samaras (winged seeds) from Pinus balfouroides, which is the paleobotanical genus-species name for the modern foxtail pine Pinus balfouriana, endemic to northern California in the Siskiyou, Scott, Salmon, Marble, Trinity, and Yolla Bolly Mountains, usually at elevations from 1,525 to 1,830 m (5000 to 6,000 feet). A disjunct subspecies of foxtail pine called Pinus austrina occurs mainly within Sequoia National Park, western slopes of the Sierra Nevada, near the headwaters of the Kings, Kern and Kaweah Rivers. Right--Samaras (winged seeds) from Pinus quinifolia. the Miocene analog of the modern Western white pine Pinus monticola, which today grows in the western United States--Washington, Montana, Idaho, Nevada, Oregon, California--and Canada (Alberta-British Columbia) at elevations from 1000 m (3,280 feet) in northern regions to 1900-3000 m (6,200 to 9,842 feet) throughout its southern reaches. |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil leaves from the 14.8 to 13.3 million year old middle Miocene Chloropagus Formation, found not far from Reno, Nevada, Great Basin Desert. Left--Salix laevigitoides, the fossil equivalent to the modern red willow Salix laevigata, which can be found today mainly from Cape Mendocino, California coast, southward into northern Baja California Mexico. It also occurs in the lower elevation western foothills of California's Sierra Nevada, southern Nevada, southwestern Utah, and central Arizona. Middle--Quercus pollardiana: A presumed extinct species of oak whose fossil leaves and acorns closely resemble today's canyon live oak Quercus chrysolepis, a common botanical component of mountainous regions in California (Sierra Nevada, Coast Ranges, Klamath Mountains, and San Gabriel Mountains, for example); also occurs in southwestern Oregon, western Nevada, Arizona, southwestern New Mexico, Baja California, and Chihuahua Mexico. Has been found growing at elevations from 500 to 2,700 m (1,600 to 8,852 feet). Right--Populus payettensis, the Miocene analog to the modern narrowleaf cottonwood Populus angustifolia, which today has a wide geographic range of natural habitat--southern Canada, southward to Montana, Idaho, Wyoming, Colorado, Utah, Arizona, New Mexico, northeast Nevada, eastern California, southeastern Oregon, western Texas, and northern Mexico--mainly along streams and creeks at elevations from 3,900 to 7,900 feet (1,200 to 2,400 m). |
 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil leaves from the 14.8 to 13.3 million year old middle Miocene Chloropagus Formation, found in the vicinity of Reno, Nevada, Great Basin Desert. Left--Betula thor: The fossil counterpart to the living paper birch (also called American white birch) Betula papyrifera, now native to almost all of Canada and most of Alaska, southward to parts of the following US states: Washington, Oregon, Idaho, Montana, North Dakota, South Dakota, Nebraska, Colorado (southernmost occurrence), Minnesota, Wisconsin, Michigan, Illinois, Iowa, Indiana, Ohio, New York, West Virginia, North Carolina, Pennsylvania, Connecticut, Massachusetts, and Rhode Island. It also occurs throughout all of Connecticut, Vermont, New Hampshire, and Maine. Middle--Cercocarpus nevadensis: The binomial term paleobotanists give to the fossil representative of today's mountain mahogany Cercocarpus betuloides, presently native to California, Oregon, Arizona, northwestern New Mexico, and Baja California Mexico, where it typically prefers chaparral communities in summer dry foothills and mountains. Right--Quercus pollardiana: A presumed extinct species of oak whose fossil leaves and acorns closely resemble today's canyon live oak Quercus chrysolepis, a common botanical component of mountainous regions in California (Sierra Nevada, Coast Ranges, Klamath Mountains, and San Gabriel Mountains, for example); also occurs in southwestern Oregon, western Nevada, Arizona, southwestern New Mexico, Baja California, and Chihuahua Mexico. Has been found growing at elevations from 500 to 2,700 m (1,600 to 8,852 feet). |
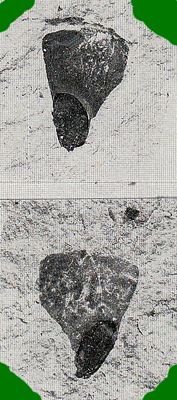 |
 |
 |
|
Click on the images for larger pictures. Photographs courtesy Daniel I. Axelrod. I edited and processed them through photoshop. Fossil plants from the 14.8 to 13.3 million year old middle Miocene Chloropagus Formation, found not far from Reno, Nevada, Great Basin Desert. Left--Winged seeds (samaras) from Abies klamathensis (Axelrod called this same genus-species from the Pyramid Formation Abies laticarpus; presumably, they are paleobotanically synonymous), the fossil genus-species of the modern Shasta red fir Abies magnifica var. shastensis which grows at elevations from 1,400–2,700 meters (4,600–8,900 ft) in southern Oregon and and northwest California (Shasta, Siskiyou, and Trinity Counties). Middle--Three cone scales with exserted bracts from what paleobotanists call Abies sherrii, the Miocene counterpart of the extant Shasta Lucia fir Abies bracteata, whose current natural habitat is confined to the Santa Lucia Mountains near Big Sur, central California coast, where it grows at elevations from 2,000 to 5,000 feet (610 to 1,520 m). Right--Leaf from Rhamnus columbiana, which is the paleobotanical name given to today's California coffeeberry (a buckthorn) Frangula californica (used to be called Rhamnus californica); presently native to California, southwestern Oregon, southern Nevada, Arizona, western New Mexico, and Baja California Mexico, preferring oak woodlands and chaparral habitats. |
|